
Neg
Innate immunity and negative vaccine efficacy.

(Long-time Unglossed readers will recognize the following as a reprisal and refinement of the innate immune suppression discussion in “Forever Spike.”1)

Is Omicron “seeking out” the Covid-vaccinated? Has Omicron finally brought “negative efficacy” out of the realm of rumor and into reality? A freshly-released study from Denmark, observing infections from November 20 to December 12 among Danes over 12, appears to offer proof:2

Previous data from the UK and Denmark have lumped all +14 days “double-dosed” into one group, or only sorted them by age, greatly obscuring the difference between those who have just received the 2nd dose and those who are a few months out.3 This new study confirms, again, that these groups are night and day in their protection against infection. Separating them allows a clear view of their disparate fates, with the result that “negative efficacy” for Omicron seems robustly affirmed for those more than 3 months past the 2nd dose. Nonetheless, the limits of these findings remain great (as discussed in footnotes4).
But rather than wage a final bet on whether the infection efficacy sky is, in fact, finally falling, the purpose of this essay is to grapple with what “negative efficacy” if true implies about the Covid-vaccinated immune response: Not “OAS,” not “ADE” as traditionally defined, but the suppression of innate immunity.
Covid Vaccines and Innate Immune Suppression
Even prior to the roll-out of the Covid vaccines, it was observable that not all individuals “exposed” to the virus, even in a household setting, went on to become infected or to score positive on a PCR or antibody test.5 This “invisible” resistance to the virus was at times referred to as “dark matter immunity” within the press. But how could some fend off the virus without even (asymptomatically) developing antibodies?
Obviously, with innate immunity.
And could the suppression of innate immunity explain why the Covid-vaccinated go on to have more symptomatic or PCR-detected reaction to the virus (if true) than an unvaccinated cohort in the same region?
Obviously, yes. For the Covid-vaccinated, this “dark matter immunity” is replaced by test-measurable infection. The result: More cases.

Possibility 1: General Immunosuppresion
Anecdotes of the Covid-vaccinated being aberrantly under the weather have become widespread since autumn, such that the simplest explanation for the suppression of innate immunity is now, “general reduced competence against viral infection.”
The Covid-vaccinated may be temporarily shielded from SARS-CoV-2 by anti-spike antibodies, but at the cost of their general immune competence against all viruses. Once sterilizing antibodies wane, increased rates of infection against the virus targeted by those antibodies must result.
A paper from May, out of the Netherlands, offers insights into the potential “rewiring” of the immune system that follows these experimental injections: Immune alarm signals against viruses and bacteria seem depressed; alarm signals against fungus oddly elevated. Here, again, the evidence comes with significant limits (again, footnotes6). What’s more, if the Covid vaccines are destructive and toxic to the body in general, a depressive effect on immunity must follow: Despite the marvels of healing of which the human body is capable, illness by default is a vicious cycle.
But perhaps you are not convinced by the anecdotes. Like me, you may not be in one of the regions where these reports of “everyone” having a cold that “won’t go away” match reality. A more satisfactory explanation for why the Covid vaccines disable innate immunity against SARS-CoV-2 only is required.
This explanation was supplied in a prediction made months ago by Geert Vanden Bossche: Residual antibodies, once they are no longer at sterilizing levels, are physically interfering with the innate immune response against the virus.7
Vanden Bossche’s theory resembles “Antibody Dependent Enhancement,” but the mechanism which he proposes is distinct from the theories that comprise that term. It is a novelty. In his telling, the residual anti-spike antibodies outcompete so-called “natural antibodies,” thereby silencing the innate immune response.
The following, however, does not represent an attempt to speak for his full version of the theory.8 It reflects where his simple, but striking premise has led my own research.
Possibility 2: Natural Antibody Silencing
Innate immunity, in the context of preventing infections from occurring in the first place, is principally the work of the Natural Killer Cells which reside directly in the tissues where our cells sit side-by-side with viruses, bacteria, and whatever else might want access. These brilliant cells (like others of the immune system) are fitted with an array of Pattern Recognition Receptors. Here, “Receptor” is a bit of a misnomer. These molecules are form-fitted to molecular shapes and chemical signatures that characterize intact host cells, destroyed host cells, and non-host visitors such as viruses; with them, the immune system is able to “see” by touch.
When the molecular milieu of the body is viewed through this lens, mere regularity often serves to mark the “pattern” of elements that deserve the immune system’s attention. Hence one of the most evolutionarily-conserved, and the first-discovered class of Pattern Recognition Receptor, the Pentraxin:
The above image depicts C-Reactive Protein, which is released by the liver in response to inflammatory signals, and assists the immune system by tagging dead cells and certain bacteria for clearance by macrophages who possess an additional receptor for CRP.
Natural Antibodies, which are potentially the evolutionary bridge between innate immunity and adaptive immunity, may have descended from Pentraxins such as this. Produced from liver-originating B-1 Cells before birth and then in the periphery throughout life (along with by some marginal B-2 Cells), they function as “free-floating” Pattern Recognition molecules, helping to extend the reach of “sensation” available to innate immune cells like Natural Killer Cells. It may be the case that adaptive “custom” antibodies are a descendent of these generic structures.
In an illuminating review by Panda, S. and Ding, J., the following image sums up everything these versatile and generic molecules are able to do.9 Note how naturally the low-affinity IgM Natural Antibody, at the top, fits to any virus with a “corona” structure:
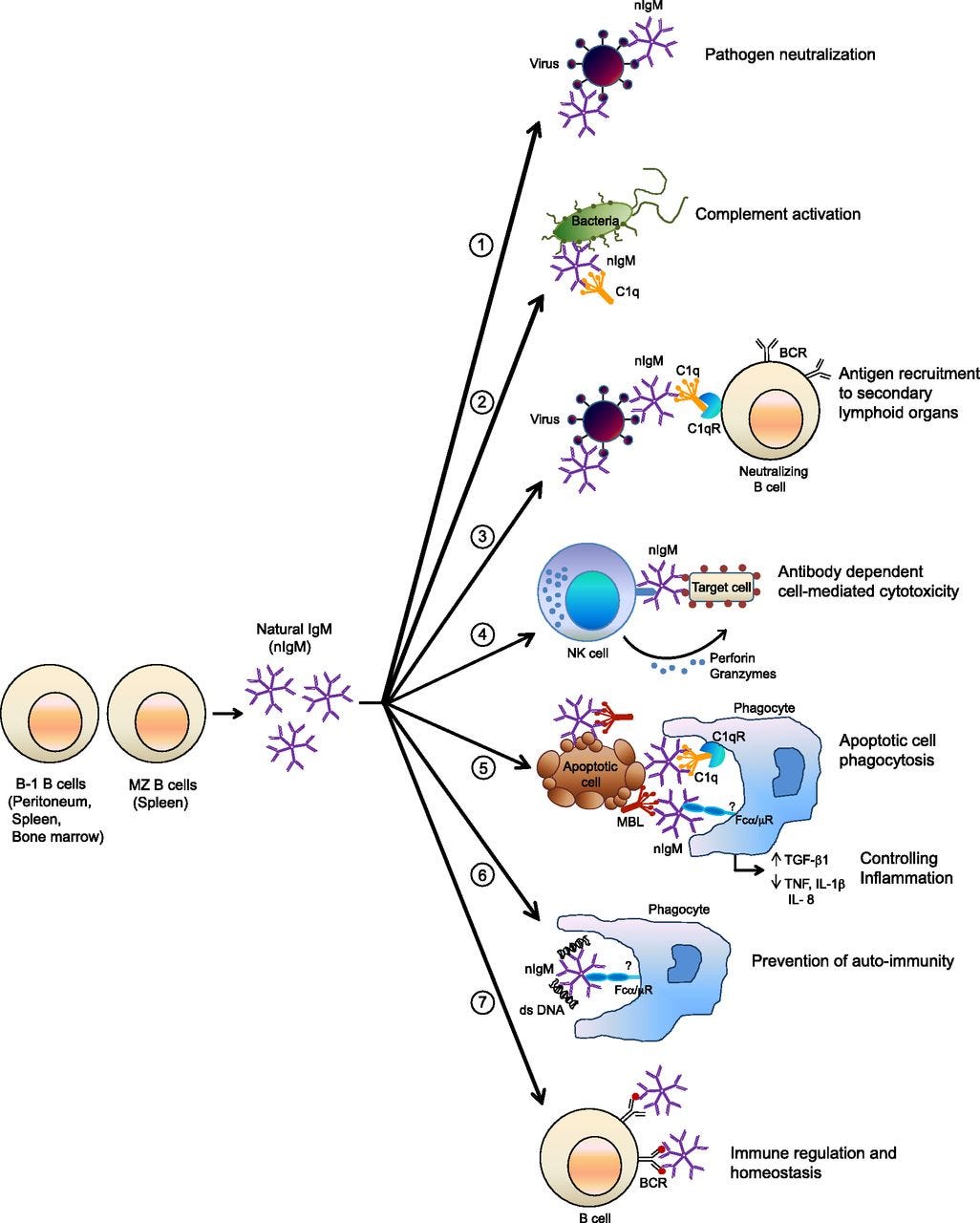
Contra the caption applied, Natural IgM probably does not “neutralize” corona-structured viruses, but only slows their entry to cells. For example, many viruses may leverage nearby bacterial molecular structures for “camouflage” as well as chemical feul, to help evade these generic defenses, as has been demonstrated with Polio Virus.10 This potentially circumvents the utility of a “one size fits all” antibody. Intuitively, Natural IgM cannot directly prevent potential infections from corona-structured viruses, or the latter would not exist. Actual clearance requires complementary activation of C1q (examples 3 and 5); again, the function of IgM is to help innate immune cells “see” the work that needs doing.
Natural IgM does, however, probably slow the progress of the virus via “general glomming,” crucially tipping the scales between virus and immune response, as well as potentially detect host cells which are expressing viral proteins on their surface and tag them for Natural Killer Cell elimination (example 4).
Natural IgM, in other words, can be thought of as the fur of the Innate Immune system. These molecules help would-be infected cells glance off light grazes; and help innate immune cells sense what is otherwise just out of reach, enhancing their competence as hunter-killers.
When the immune system creates custom antibodies against any particular virus, this naive competence is sacrificed; innate immunity is shaved raw. Custom antibodies bind to the virus antigens with higher affinity than Natural IgM, potentially leaving Natural Killer Cells cells blinded. No wonder modern medicine’s fixation on detectable serum antibody - induced via vaccination - has been a perpetual, Quixotic campaign targeting the wrong dragons. There is so much more to immunity than just antibodies; and proper enrollment of all elements depends on the Antigen Presenting Cells that normally perceive the original encounter via the same Pattern Recognition Receptors mentioned above. Remove the pattern - natural infection - and you remove the proper response.
Handing over immune response from innate to adaptive immunity, in the wake of an infection, implies that cellular immunity - the defenders where our cells sit side-by-side with microbes - must be retrained to recognize the molecular signatures targeted by new antibodies directly. The Natural Killer Cells so adept at protecting against unfamiliar challenges must be supplemented by Resident T Cells proficient in recognizing the molecules that appear alongside of and even before those targeted by antibodies.11 Significantly, the adaptive immune system tethers the refinement of long-lasting IgA and IgG antibodies to T Cell mediation:
Adaptive immune response
Antigen-Presenting Cells migrate to Secondary Lymphoid Organs and present antigen to: B Cells and T Cells
T Cells: Helper T Cells respond to APC instructions to orchestrate downstream B Cell, Killer T Cell, and Resident T Cell responses
B Cells:
Non-mediated (“TI,” or T Cell Independent):
Short term IgM and Short-Lived Plasma Cell response - putting out the fire of viremia
Germ Center T Cell mediation:
Somatic Hyper-mutation for high-affinity IgA/IgG refinement, leading to:
Long term IgA/IgG and Long-Lived Plasma Cell response
This elaborate gate-keeping serves to guard against long-term autoimmune B Cell responses, and to ensure that long-term antiviral B Cell responses are accompanied by robust cellular immunity. Any B Cell response that is going to last a long while, lest it lead to immune dysfunction, should be closely regulated by the T Cells which received their instructions in turn from Antigen Presenting Cells. Again, this process is sabotaged by vaccination, which seeks explicitly to author the response in a given direction.12
In the wake of a natural infection with SARS-CoV-2, for example, some of the memory T Cells that are activated by Helper T Cells follow a cascade of inflammatory signals to home in on the respiratory and gastro-intestinal tracts where the virus is active. There they will reside permanently, recognizing the signatures of the virus or its replication in cells, should they reappear.
Gradually, as antibodies fade, this lifelong T Cell recognition compensates for the disadvantage of residual custom antibody binding to the virus which blinds Natural Killer Cell recognition. With enough antibody fade, both innate and adaptive cellular immunity become functionally competent at suppressing later challenge by the virus, resulting in durable immunity (with B Cells available to ramp up antibodies to guard against viral spread into the bloodstream).
In the wake of Covid-vaccination, no migration of Resident T Cells takes place, and the innate immune system is left at the mercy of the virus due to the silencing of Natural IgM.13 Once anti-spike antibodies fade below the boundary of “sterilizing,” but still out-compete Natural IgM, infection must result.
Hence, negative efficacy.14

Further reading:
“Burned All My Notebooks.” (Brian’s crackpot Immune Equilibrium theory.)
“Crackpot Corner: Marekspocalypse Edition.” (More on “onboarded” viruses.)
“(Not) Coming Up.” (OAS is not real.)

See “Forever Spike,” which besides innate immune suppression also discusses “vaccine escape” and “Marek’s Effect” (rather dismissively).
The secret to growth on Substack, as far as I can tell, is the same as Twitter, Instagram, etc: To endlessly repost previous popular posts almost word for word, prompting newer readers to share them so that even newer readers will arrive who can be spammed with the next previously popular repost (minimum number of previously popular posts needed: 2).
Whether this practice wastes the time of early subscribers - the readers who provided support earlier, and obviously have already seen the content being republished - is irrelevant; it’s not like these people can “subscribe again,” after all. Those alleged “supporters” are therefor irrelevant as all they do is unfairly make the author have to provide more than two worthwhile thoughts in order to grow.
Comparing Omicron (distinguished in readily-available Delta-targeted PCRs by its vintage 452L gene) case rates to non-Omicron, additionally, may distort the picture: If the Covid vaccines still work well against Delta for a few months after dosage, Covid-vaccinated case rates for Delta will be less than for the unvaccinated. This seems to easily explain the notorious report released by Statens Serum Institut in Denmark (https://www.ssi.dk/-/media/cdn/files/covid19/omikron/statusrapport/rapport-omikronvarianten-18122021-wj25.pdf). Nonetheless, the Covid-vaccinated seem slightly over-represented. I fashioned my own crude guess for the “not vaccinated” rate among Danes 12 and older:
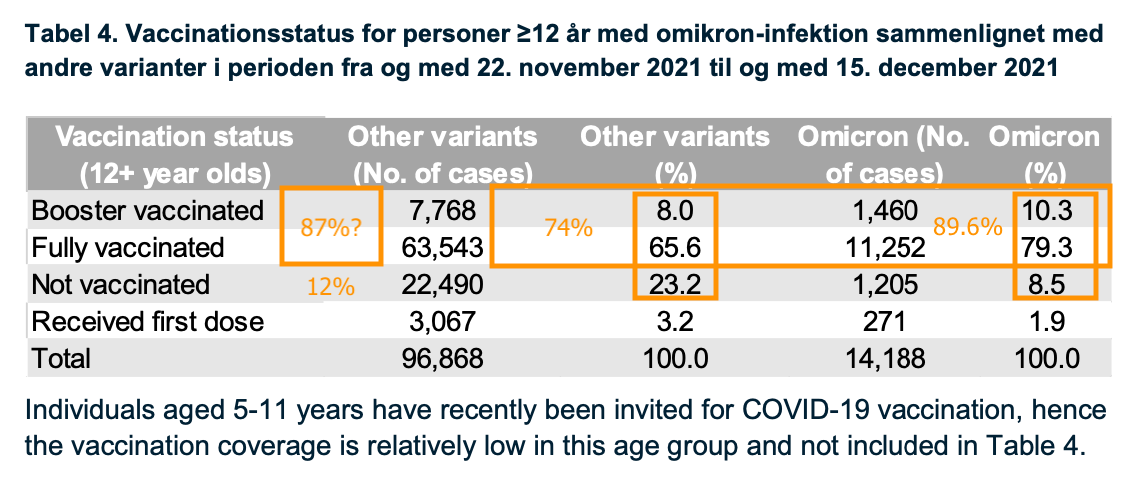
A more direct demonstration of how infection efficacy against “other variants” can create the illusion of negative efficacy for Omicron is possible using the unvaccinated+infected alone. For these:
Unvaccinated+infected real Omicron rate
1,205 / (22,490 + 1,205) = 5.1%
Unvaccinated+infected if 50% “other variants efficacy” Omicron rate
1,205 / ((22,490 x .5) + 1,205) = 9.7%
Unvaccinated+infected if 80% “other variants efficacy” Omicron rate
1,205 / ((22,490 x .5) + 1,205) = 21.2%
Mathematically-applied efficacy against other variants would give the unvaccinated + infected “4 times” the rate of Omicron they had before.
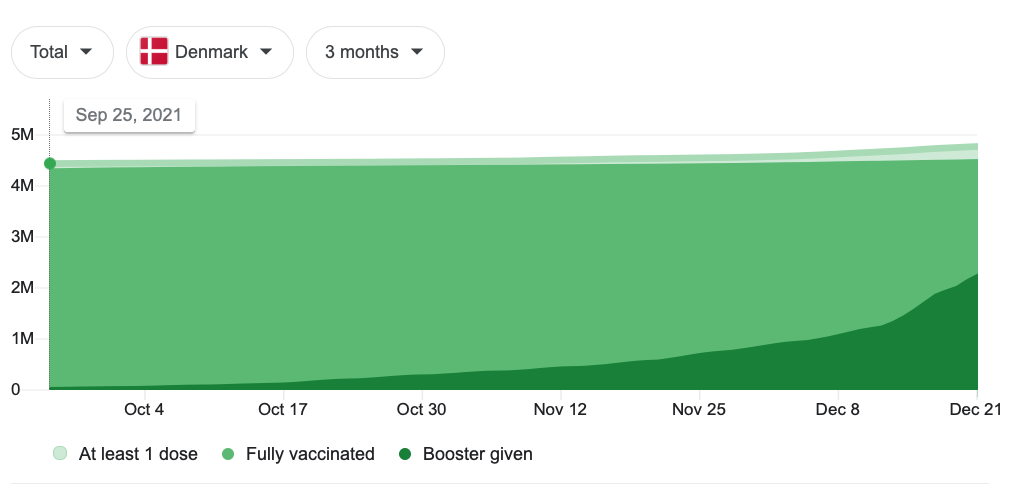
Among which: The authors do not provide their calculated exposure “person-days,” nor the value for cases among the unvaccinated during the November 20 to December 12 observation window (again, only Danes 12 and over are observed, to reduce problems with a changing denominator for the unvaccinated). These results are further distorted by the authors’ undisclosed adjustments for age, sex, and geographic region. Note that the 91-150 day efficacy values for Delta are patently implausible. (Edit, Dec 27: I am a dolt. These values are for to-date infection efficacy, not “real-time” efficacy (such as in the infamous UKHSA charts); in a to-date view the vaccines will still statistically benefit from “prevented” (really, just delayed) infections until a big wave passes through. Since Omicron infections only started in November, the to-date view is essentially a real-time view.)
Back to the main findings RE Omicron, just as with the autumn data from the UK, there is a potential that the 91-150 day cohort is playing “catch-up” with the unvaccinated (less plausible, in this case, since there was no preceding wave before the observation window). Or, negative efficacy could be a statistical artifact of the high uptake of boosting prompted by the recent wave, which potentially selects “uninfected” out of the “fully vaccinated” denominator. Things are in fact quite messy on the ground:
Scare-quotes here as a nod to the unclear role of sick-to-well transmission in SARS-CoV-2 spread.
Nonetheless, household contact appears to result in an immune response even for those who do not develop symptoms or antibodies. This potential glimpse into innate suppression was recently supplied by Swadling, L. et al. “Pre-existing polymerase-specific T cells expand in abortive seronegative SARS-CoV-2.” Nature. 2021 Nov 10.
Here, close encounters with the virus leave evidence of alerted T Cells against the generic coronavirus polymerase, the first protein which is created and presumably available for inspection when SARS-CoV-2 enters a cell (for further description of the coronavirus replication process, see “Doppelgänger”). This may be the sign of successful suppression of T Cells, or the T Cells may merely be alerted by the aftermath of suppression by Natural Killer Cells. In either case, it provides a glimpse into the “dark matter” of exposure without antibodies or symptoms (and suggests prior familiarity with coronavirus either helps or does not hurt immunity against the virus, impossible if “OAS” were meaningfully true).
See Föhse, F. et al. “The BNT162b2 mRNA vaccine against SARS-CoV-2 reprograms both adaptive and innate immune responses.”
Limits: Many relevant markers show drift in both directions (half of subjects reducing and half increasing their immune response after the injections). There is no apparently known baseline for the normal rate at which the measured cytokines for inflammation and tolerance “drift” higher and lower, absent the injections. Further, the authors fail to include any non-Covid-vaccinated controls in the study. As all the test subjects are healthcare workers at the Radboudumc Nijmegen, there may have been environmental factors (such as high bacterial challenge) leading to synchronized immune sensitization or tolerance at the beginning and end of the local Covid vaccine rollout.
Nonetheless, the paper is more extensively reviewed by Jessica Rose in an essay by the same name, which is a must-read for its “fictional” account of an encounter between SARS-CoV-2 and the immune system:

In this model, Omicron, with its more alien spike protein structure, accelerates the arrival of this transition. But note that the “General Immunosuppresion” proposal is also supported by the earlier transition. In either case, the boundary between “sterilizing” and not, against Omicron, becomes much higher than for previous variants - and thus is crossed earlier on as antibodies fade after the 2nd (and presumably 3rd) dose.
For a recent dissertation, see Vanden Bossche, Geert. “Omicron: The calm before the tsunami?” (2021, December 9.) Voices for Science and Solidarity.
Panda, S. Ding, J. (2015.) “Natural Antibodies Bridge Innate and Adaptive Immunity.” The Journal of Immunology. 2015 Jan 1;194(1):13-20
See Kuss, S. et al. (2011.) “Intestinal microbiota promote enteric virus replication and systemic pathogenesis.”
See footnote 5.
In the case of the Covid vaccines:
The ideal spectrum of epitopes for antibody targeting cannot be created; the immune system must focus exclusively on the spike and may target host-homologous epitopes that otherwise would not have been “of notice,” leading to autoimmune antibodies. The spike protein is a smorgasbord of such potential epitopes.
Long-lived plasma cells may infiltrate sites of inflammation caused by mRNA transfection and spike protein expression. Here, not only will they be less useful against later viral challenge in the respiratory tract, they may also contribute to long term autoimmunity.
As long-term B Cell responses are refined, anti-spike antibodies may be class-switched into IgE, which can potentially infiltrate tissues and cause a later inflammatory response, leading to Antibody Dependent Enhancement. Vaccine developers have essentially no way to measure whether this is the case and therefor do not attempt to do so. See “Lurking Leviathans.”
Note, again, that this does not represent Vanden Bossche’s model of the problem (footnote 8). In particular, he is fixated on “trained” Natural Antibodies, which does not seem supported by any of the research. As he proposes that adults have stronger, “trained” Natural Antibodies, I can’t really understand how his version of the model accounts for the obvious innate immunity of children against the virus. Additionally, he appears to propose that Omicron is sufficiently distinct in spike design that Natural Antibodies can now be “trained” on it. Again, I don’t understand how this accounts for negative efficacy against Omicron (implying that low-affinity, non-”sterilizing” anti-spike antibodies are still outcompeting Natural Antibodies). On the other hand, it’s only recently that I’ve come around to the idea that Natural Antibodies play a strong role in innate immunity at all, so my record on intuitively disagreeing with vanden Bossche is not holding strong.
The question being, as ever, whether “breakthrough” infection results in a compensatory proliferation of competent T Cell response against the broad spectrum of epitopes for SARS-CoV-2, and migration of Resident T Cells to the respiratory and GI tract. I remain optimistic (see the brief review of reinfection results by Goldberg et al. in “Darmok and the Spike Protein at Tanagra”).
Thanks for your well-reasoned take on this, and for a more helpful explanation of how "natural antibodies" work than Geert has ever offered.
Although I think your two possible explanations probably rank higher at this point, I would like to offer a third that may rise in relevance as more people get 3, 4, or more injections of the same genetic spike vaccines.
Specific Spike Tolerance
Foreign proteins can function as both antigens (generating an antibody and inflammatory response) and tolerogens (generating a tolerance response that inhibits inflammation). We are constantly exposed to foreign proteins that are not associated with pathogens, and when our immune system treats them as antigens the result is an allergic reaction.
Allergic reactions can often be treated by regular controlled injections of the offending protein, which activates tolerance mechanisms. I had this experience personally after developing an allergy to bee stings, which was successfully treated by allergy shots.
My concern is that the mechanism of delivery of the spike protein in genetic vaccination does not "look" to the immune system like a viral infection, and also that the ensuing antigenic response and attack on spike beating cells "looks" like an autoimmune reaction.
One expected outcome of this, which would be exacerbated by boosters, would be the activation of tolerance pathways that counteract and ultimately downregulate the initial immune response to spike protein, even as antibody levels remain high and receive a boost upon further injections.
The cells to keep an eye on here are regulatory T cells (Tregs or FoxP3+), which are well studied in cancer research (because they can prevent the immune system from recognizing and destroying metaplastic cells) but so far not much examined in the context of SARS-CoV2 vaccination.
Given the freshly jabbed are
1. not sequestered from society
2. probably jabbed in high viral load/dose traffic locales
3. human, and unlikely to retain the "I am not vaccinated [sic] until 14 days time" info in their "jabbed = freedom" minds...
I would seriously love to see the stats of that 1-14 day post jab VE rate and the impact in general on cases, etc. It almost feels like cheating to exclude that period of time.