
Burned All My Notebooks
Immune Equilibrium, three ways.

[Warning: Draft. I will be editing this significantly in the coming weeks. The first and third segments, in particular, require clean-up. However, as all the relevant thoughts are now on the page, I am making the essay available for any readers who are interested, despite what must be several dozen half-edited sentences which contain tense disagreements etc.]

Burned all my notebooks, what good are notebooks?!
They won't help me survive.
My chest is aching, burns like a furnace!
The burning keeps me alive.1
(Preceded by: Preface.)

Equilibrium
Viruses are a part of us, and vice-versa.
There is no meaningful distinction between the endogenous and viral, exogenous component of the genome of humans or other species; together they make up the super-genome for every organism or colony of that species.2 All species-specific, endogenous genomic elements whose phenotypes are “exploitable” by viruses are equally validly explained as elements designed to better sustain a rich virome, improving the odds of competent horizontal gene transfer in general and enabling calibration of likelihood of horizontal gene transfer according to environmental and stress stimuli. Thus, the immune system of any given species for which there is a virome (likely all multicellular life at least) does not and cannot favor the survival of one component of the super-genome - endogenous or viral - over the other.
Prokaryotic colonies and multicellular organisms are both designed to, upon development / birth, onboard multiple environmentally-encountered, species-compatible, and immunity evading viruses with built-in dormancy timers that ensure survival of the host colony / organism. These onboarded viruses provide a reservoir within the viral portion of the genome to insulate against depopulation shocks; it is plausible that the genome possesses competence at assigning an affinity for this portion of the genome to resiliency-conferring genes, be they manifested in the form of biofilms or anti-sociality, and altruism-conferring genes, be they manifested in the form of plasmids or pro-sociality - plausibly favoring the preservation and stability of these genes even above the endogenous genome’s protein script set. “Eternal spread” is not necessary to preserve Onboarded Viruses; these viruses therefor are not efficient at evading antibodies and thus only create symptomatic illness in the young.3
Multicellular organisms additionally possess fluency with high-efficiency, dormancy-deficient viruses which occupy the receptor-mediated endocytosis infection bandwidth, based on staggered vulnerability and promotion of “eternal spread.” These viruses do benefit from antibody escape, and therefor continually re-challenge the immune system with the task of repelling later infection. Achieving and maintaining competence in this fluency is merely the “external” element of the immune system’s bivalent menu of hazard, the other being “internal” - cancer and autoimmune disorder - neither side is intrinsically more hazardous.4 Thus, core immune competency and fluency with eternal spread are essentially interchangeable.
Moreover, “viruses” as we think of them are merely the exogenous, social element of both the super-genome and the embryonic and post-embryonic gene expression operating system for a given species, the endogenous portion of which includes myriad genes which but for their retention in the nucleus are otherwise indistinguishable from their exogenous counterpart - and in fact enter a transitional state during embryogenesis. Therefore, a new model of genetics is required which acknowledges the (Social or) Collaborative Genome.5 The endogenous genomes of multicellular organisms pass through time in a constant flux between stasis/evaluation (fixed state) and election/revolt (embryogenesis).
Lastly and most significantly, the multicellular immune system is designed to switch off during times of extreme stress, to facilitate genomic innovation during embryogenesis. The genome of a given species is best understood as a society of genes of different temperaments, whether “selfish” or passive, which are capable of authentically sensing and evaluating for themselves whether the current social order is working or not, and of mounting a social revolt in the latter case. This “revolt” is experienced as increased susceptibility to infections, which grant novel, viral genetic code access to the germ cell as a last ditch bid for survival of the genes which compromise the society, thus explaining the prevalence, if not near-ubiquity of endogenous retroviruses in the genome. This entirely accounts for the advantage of and desire for the phenotypic proliferation of exogenous genes - viruses - from the perspective of endogenous genes, once again affirming the existence and essentiality of the super-genome.
There is, and likely never will be, any better mechanistic explanation for evolutionary leaps and for the generation of pro-social neurological traits. Genomic Revolt, in fact, makes both those things easy to explain.

The first justification for Immune Equilibrium theory was already presented in “Die Herd,” but is repeated below in an updated form.6 However, I have not copied over the list of predictions for Immune Equilibrium included in the footnotes of that essay.

Immune Equilibrium, and the Impossibility of Herd Immunity
Immune tolerance of bacteria is the essence of “immunity” from bacteria.
Bacteria are directly and indirectly critical to most or all facets of immune competence.
For viruses, the immune system’s “goal” is to balance host and viral survival. This is why the immune system acts like that is its goal. Antibody levels are meant to drop after a cellular immunity rest period, as a bid for reencounter and re-spread with a given viral challenge.
This strategy removes selection pressure for core immune-evading traits within respiratory and gut viruses, preventing the immune system from having to “solve” the unsolvable in order to ensure basic survival; it additionally removes pressure for antibody escape, slowing the generation of novel antigenic structures in any given virus.
Further:
If a virus happens to take on core immune evasion traits, the fitness gains are either contextual (for rare-frequency spread pathways, immune evasion is probably essential to survival) or short-lived (elimination of host species). The virus must learn to turn off replication and enter lysogeny / dormancy immediately upon access to vital organs, analogous to how bacterial viruses (phages) turn off replication at a certain threshold within a given colony. This explains timed dormancy in common childhood viruses like herpes viruses and hypothetically hepatitis viruses. These viruses are not “defeated” by the immune system - they turn off themselves. Reactivation of dormant, “onboarded” immune-evading viruses is plausibly a reply to tissue stress sensors that indicate a decline in host health.7
Dormancy-triggering timers are plausibly calibrated to the immune competence of their host species and may become overclocked over time as a result of artificial immunization (Marek’s Disease herpesvirus, possibly Measles and hepatitis viruses).8
For viruses in high-frequency spread pathways (respiratory and gut viruses) without immune-evading traits, there is no evolutionary curve to “more contagious / less deadly.” Novel respiratory viruses without core immune-evading traits merely become another familiar, host-species-sustained virus immediately after first encounter. Contagiousness is thereafter defined by the portion of immune bandwidth a given virus happens to occupy (such as the incredibly versatile ACE-2 receptor pathway). “Deadliness” is thereafter defined by the immune competence of the host population at a given moment. Such respiratory / gut viruses may in fact feature legacy evasion / dormancy genes, but these will be rarely triggered: As stated above, the immune system’s promotion of reencounter and re-spread removes any incentive for these viruses to sabotage cellular immunity.
“Herd immunity” to almost any virus via widespread, persistent “sterilizing immunity” is thus not naturally possible, regardless of animal reservoirs.
“Die Herd” extensively caveats that this argument and Immune Equilibrium itself are, to a great degree, merely putting a name on a movement in modern research that has been underway for decades, but which appears reluctant to name itself.
On the matter of acknowledging the obvious significance of the discovery of what this essay goes on to name as the Transposon Cycle, there have been a handful of hits just to the edge of the bullseye; the closest, I believe, came in 2016.9 So in that sense, I do not claim authorship. In fact, I feel that with this as with many other things, the collective consciousness seems to be arriving at the same truths simultaneously throughout the last half of 2021, as if modern medicine, by perpetually running its flawed programming through the mind of outside observers over and over and over, is finally having all of its critical errors recognized and voiced aloud.10 Thus, it was no surprise that the above version of the argument emerged almost simultaneously in an essay by another writer, though without the emphasis on immune-evading virus dormancy-timer competence.11 And of course for all I know, this entire theory is already the subject of popular Tik-Tok videos in Croatia.
The argument for Tenet 3 of Immune Equilibrium above is reverse-teleological: Since we wouldn’t be around if things were otherwise, the immune system’s goal must be to promote survival of viruses; therefor the immune system does promote the survival of viruses.
This essay, however, forwards two additional and entwined arguments for Tenet 3. The first is an observational argument, which itself advances the first part of a “new” model for genetics overall. The other a second reverse-teleological argument for Tenet 3, which is even stronger than the argument above, and completes the “new” genetic model. The former is a dense and sweeping construction, though shallow; the latter should render the entire theory obvious, if not impossible to unsee.
Part 2, to be posted later, will attempt to frame Immune Equilibrium in terms of the history of the research. As the microbiome’s inevitability, ubiquity, and benefits have already been rendered more or less orthodox by that same research, Tenets 1 and 2 will be elaborated there. Part 2 will also offer comments about seasonality and plagues, more or less the only two subjects wherein Germ Theory has not already proven deficient.
Readers of all types are hereby warned that the author is not fluent in molecular and genomic biology - throughout my rushed construction of this theory I have considered the matter of what happens on the chemical level of the genome unimportant, and what happens in the chromosomal level to be merely a hint of a larger pattern.12 Hence why genetics was already close to unveiling this pattern in the cytological era of the 1930s and 40s, only to wander in the wilderness upon the detour into molecular description.
In fact, of the preparation I have conducted over the last months, more time has been spent looking for any realms of biological science whose current understandings could in any way contradict the Immune Equilibrium model, than confirming the existence of new findings that support it (“new” meaning within the last ten years, since as mentioned previously, the theory was intuited ten years ago from just a cursory review of contemporary research, and incorrectly taken as likely to arrive without any work on my part).13 My research, in other words, sought to run Immune Equilibrium through the same century of findings which have so thoroughly dismantled Germ Theory, not to “figure out” the former. Nonetheless, nearly all subsequent findings did go on to support and elaborate that dim intuition, establishing that the apparent evidence for Immune Equilibrium is everywhere one can look.
But it nonetheless remains that I am as imperfect a messenger as the theory could probably get. Certain terms may therefor be completely mis-applied; certain facts grossly mangled. The overall claim will, I wager, still stand.
See me put things together
Put them back where they belong
When I look at each other
Have I always been singing the same song?14
In a Perfect World
Terms:
Prokaryote World
Prokaryote: A unicellular organism without a nucleus, genetically distinguished into bacteria or archaea. “Prokaryotic” in this essay will typically refer to a quality that has been observed in bacteria. No multicellular organisms are prokaryotes, but many single celled organisms are eukaryotes. This essay limits its discussion to prokaryotes (single cells), which form colonies, and multicellular eukaryotic cells, which, of course, form multicellular organisms.
Phage: A virus that resides in an active bacterial genome.
Lysogeny: The integration of a virus within a cell, resulting in dormancy.
Dormancy, or latency: The cessation of viral replication within a cell, as a result of lysogeny. Dormant viruses may either reside in the cytoplasm or take up residence in the DNA of the cell. Dormant viruses may later re-activate, leading to (resumption of) replication and shedding. Dormancy-capable viruses thus appear to be in possession of signal/timer logical pathways of high sophistication.
Biofilms: Structural collections of bacterial cells bound by natural polymers expressed by those or previous bacterium. This essay will also use “biofilm” to refer to the natural polymers themselves.
Spore: As in endospore. A hibernating bacteria which has encased its core in a protective shell, potentially remaining viable for eons.
Multicellular World
Embryogenesis: The development of the multicellular embryo, during which stem cells are formed and programmed according to their appropriate fates and descendent cells are differentiated and instructed.
Transposable element: Also, “transposon.” A gene which is capable of replicating itself within the genome. These are the most prominent type of “selfish” genes - genes which appear to prioritize their own duplication within a genome over the survival of the genome as a whole. Transposable elements are unlocked during embryogenesis, indicating a role in conducting cell differentiation that involves sophisticated and interconnected viral signal/timer logical pathways, rather than the mere reading of static genes as an “instruction set.”
Endogenous retrovirus: A transposable element which has been identified to share genes with a wild virus.
Prospective endogenous retrovirus: A virus that has encoded genes into the germ cell (sperm or egg) of its host. These genes will be incorporated into the endogenous genome of offspring (if they do not render the offspring inviable), becoming a static endogenous retrovirus. (Viruses are not capable of transitioning from the viral genome to the endogenous genome without access a germ cell that goes on to produce viable offspring of the host.)
Onboarded Virus: A dormancy-timer virus with immune-evading traits, typically incorporated into multicellular organisms during early life, that may reactivate in response to signals. Onboarded Viruses constitute a segment of the super-genome that is both stable and nearly ubiquitous, yet does not reside within the endogenous genome, though it may participate in “endogenous gene expression.” Thus, Onboarded Viruses most closely resemble temperate phages.
Genome World
Gene: Any set of information which both is encoded in the format compatible with RNA-based biomolecular machinery, and exists in a frame of reference within “cellular time”: It has arrived to the present from a previous cell which no longer exists, and is contextually capable of perpetuating into future cells after the current cell no longer exists. The particular quality of all sets of information that meet that condition, is realizable self-interest, whether or not the given set of information is capable of or temperamentally inclined to promote that self-interest.15
Genome, traditional: A host of genes which are collectively “active,” as in engaged or potentially available for collective gene expression, at a given moment.
Gene expression: The engagement of genes with the extra-genetic, RNA-based biomolecular machine. Gene expression, traditionally defined, is the “reading” of genes to produce biomolecular outputs. In light of transposable elements, gene expression also authors genetic inputs based on feedback from the biomolecular machine, violating the boundary between the biomolecular machine and the genome. Moreover, multiple realms of “high-intelligence gene expression” have recently been found to involve DNA demethylation, implying the unlocking of transposable elements and viral signal/timer logical pathways.
Phenotype: Any quality induced by gene expression which, in a material context, creates a distinctive outcome. This ranges from the creation of proteins to hair color. Additionally, an individual organism which is contextually distinct from other organisms.
Genome, novel: Any set of genes whose endogenous share depends for perpetuation on a particular individual biomolecular machine, and whose viral share depends for perpetuation to some extent on future instances of biomolecular machines of that type.
Super-genome: An overall genome as comprised of both endogenous and viral shares of genes. In this essay, the definition of “super-genome” is the same as “genome,” including in the novel definition above; the former will be employed whenever an explicit inclusion of the viral share of the genome is appropriate.
Collaborative Genome: The viral share of the genome. As opposed to endogenous genes, the genes of the Collaborative Genome may be employed and edited by other colonies / organisms / species. Within the super-genome, the Collaborative Genome is both an ideal location for contextually valuable genes that need to be conserved in timespans longer than a few generations, and for experimental genes. This is true for both prokaryotic and multicellular super-genomes.
Genomic society: Also, “The society of genes.” Yet another way of describing the super-genome. The super-genome may be thought of as a society of genes, with their own self-interest and set roles, etc. These phrasings will be used sparingly, when they are illustrative.
DNA demethylation: Who knows. It’s a thing that does things and then a bunch of stuff happens and it’s transposon time.
Transposon Cycle: The alternation between stasis/evaluation (fixed state) and election/revolt (embryogenesis), as experienced by the genome.
Immune Equilibrium: An umbrella term for all material manifestations of the promotion of bacterial and viral survival within the multicellular endogenous genome; and within both prokaryotic and multicellular super-genomes, the promotion of viral feature-storage and innovation by the endogenous genome and the promotion of endogenous genome survival by the viral genome.
Precepts and Conclusions:
Bacteria use plasmids to share beneficial genes, including genes which assist in genome repair. Plasmids are non-destructive to the donor, but require metabolic energy and chemical resources.
The ideal design for an efficient plasmid is one that consumes all the available energy and resources of the host. This is a virus. (Bacterial viruses are described as “phages”).
Phages confer and contain not only directly beneficial genes but assorted sensors and timers. Sensors are genomic algorithms which infer the current status of the bacterial genome from the rate of encounter with chemical signals emitted by both naked and phage-enhanced members of the same host genome. To be crude, one may imagine a blind and deaf dog, which nonetheless understands at every moment whether it is amongst kin and whether the pack is content or under stress. Sensors may be used both to evaluate the status of the genomes of neighbor hosts and to prompt communication with them - to bid for plasmids, for example. Timers are used to prompt dormancy - the incorporation of phages into the host cell. Phages principally achieve fitness by mastering the ability to calibrate dormancy timers according to their host genome. As even singular quorum-sensing molecule pathways have been associated with observable bacterial collaboration, the potential environmental awareness and “intelligence” of phage-enhanced bacteria with access to multiple signaling pathways is significant.
Dormancy-timer viruses are likely the most intelligent and sophisticated of all genes in either the prokaryotic or multicellular genome. Therefor, bacteria are genetically just as sophisticated as multicellular organisms. Therefor, dormancy-timer viruses are plausibly responsible for the facilitation of multicellular life, even without any mechanistic explanation.
To reiterate 3 and 4, the activity of dormancy timer viruses enables the phenotypic manifestation of bacterial temperament. Bacteria enhanced by phages are capable of alternating between modes of contentment/generosity and resilience/stringency, depending on the friendliness of their environment, as detected by genomic sensors. These genomic alternations are manifested phenotypically as biofilms, spore-conversion-reversion, and alteration of metabolic diet. Phage-enhanced bacteria thus commonly proliferate in harsher conditions. In this way, the exogenous, viral genome “leads” their genomic society through rough times and ensuring long-term survival.
The achievement and perpetual perfection of the sophisticated gene intelligence within the prokaryotic super-genome is intuitive when considering the regularity of cataclysmic events confronted by the bacterial genome. For context: To bacteria, the evaporation of a puddle is a cataclysmic event.
To elaborate 5 and 6, the bacterial genome is not “invested” in the survival of the host cell. The bacterial genome is a society of genes. The regularity of cataclysmic events confronted by the bacterial genome determines that an arbitrary, exaggerated rare survival rate of 1/1,000,000 to cataclysmic events is worth an (arbitrary, exaggerated) host mortality rate of 1/10 via viral shedding, i.e. infection of bacteria by viruses not in the dormancy phase of their signal/timer pathway. In fact, only such a scheme as this can explain the successful conquest of Earth by organic matter.
Thus, viruses are again responsible for the (in this case, circumstance which could make possible) proliferation of multicellular life.
Multicellular endogenous genomes consist either heavily or predominately of “selfish” genes, i.e. transposable elements.
Transposable elements and their function were discovered by Barbara McClintock in 1944-1950, foreshadowing the later discoveries which set the ground for Precepts 1-8, as well as every discovery in the realm of genetic expression in the last 75 years. Transposable elements up-regulate their activity during embryogenesis in response to damage to the genome, leading to a rate of mutation among progeny which, as observed by McClintock, had never been observed in any experiment prior to hers.
Thus, the endogenous portion of the multicellular genome is able to sense crises/cataclysm as well. The sophisticated sensor/timer skill-set of the prokaryotic viral genome and the fitness advantage conferred by “selfish” genes during cataclysmic events not only perseveres in the multicellular realm; it has been made essential to it.
Thus, there is no valid distinction between transposable elements of the endogenous multicellular genome, and viruses. They are equally “selfish” members of the super-genome of that organism which nonetheless have developed contextual self-regulation (there is no higher “author” to promote transposable element replication in the McClintock seed experiment, thus there is no “author” that normally represses it; transposable elements, like bacterial viruses, repress themselves).
Viruses frequently transition from the Collaborative/viral to the endogenous portion of the genome. Many (endogenous) transposable elements are already recognized as “endogenous retroviruses” based on overlap with recognized viral genes (and a lack of recognition does not imply a lack of ancient viral origin). It is plausible that all transposable elements are in fact ancient viruses which now reside in the endogenous genome; that any given super-genome is predominantly a vessel in which a society of viral genes resides, that the endogenous portion of the genome is no more productive/participative during embryogenesis than the Collaborative portion, and that the endogenous genome facilitates the generation of proto-viruses.
Transposable elements are suppressed by DNA methylation. DNA is demethylated during embryogenesis, “unlocking” transposable elements. Paternal and maternal DNA are fully demethylated a few days after fertilization, and partially remethylate afterward. In mice:
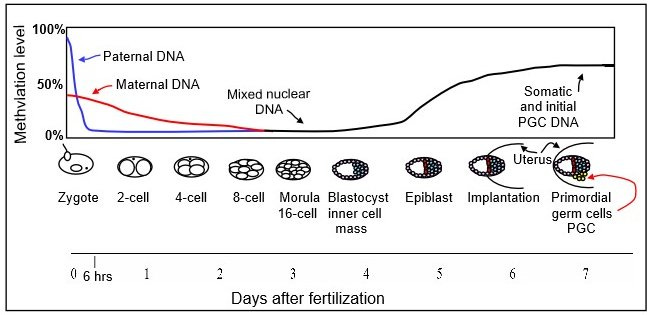
Source: “Chaya 5260” Recent research has demonstrated that endogenous transposable elements generate viral particles within the multicellular embryo.
Thus, transposable elements are plausibly critical to not just the achievement but the possibility of the achievement of embryogenesis, and therefor multicellular life itself. (Non-endogenous) viruses themselves have been discovered to be present in fertilization in some form, and may also participate in embryogenesis. Just as phages seem to regulate bacterial colonies according to signal/timer logical pathways, transposable elements seem to conduct embryogenesis according to contextual logic; implying that embryogenetic logic is viral logic.
This implies that embryogenesis is not the product of a “script” within the endogenous genome. It is a collaborative, but distributed effort between genes to interpret, reply to, and set in place a dynamic range of biomolecular conditions. This mode of embryogenesis may be described as “election,” when genes within the genome determine their coming roles in the fixed portion of the Transposon Cycle. As with human societies, the typical result of an embryonic election is that nothing changes (embryogenesis, again, only begins after the classical gene selection events of meiosis and fertilization; but it may feature the participation of non-endogenous viruses in some manner).
A prospective endogenous retrovirus - one that is dormant in the germ cell before fertilization, may disrupt embryogenesis, rendering the host’s endogenous genome essentially defective, or may be compatible, resulting in new static endogenous retrovirus in the genome. The resulting endogenous retrovirus may confer a beneficial phenotype, or be neutral.16
Embryogenesis is also plausibly indirectly mediated by Onboarded Viruses in many, if not all forms of multicellular life. To focus narrowly on mammals, reactivation of dormant viruses adopted in childhood could prompt activation of maternal hormonal pathways during pregnancy that are responsible for both prosocial and antisocial psychological outcomes, as well as other phenotypic responses to lifetime stress patterns. For other multicellular organisms, Onboarded Viruses could potentially mediate the hormonal programming of egg yolk, endospores, or for insect colonies, the psychological programming of phenotypes.
Onboarded Viruses are an ideal portion of the genome in which to move stable genes that need to be preserved over possible successive generations of counter-signaling - which, in other words, need to be “insulated” from natural selection, to ensure that sudden changes in external conditions do not render the super-genome unfit. Thus, just as phages confer the classic traits of resilience to bacteria - biofilms, metabolic variation - Immune Equilibrium predicts that Onboarded Viruses contain genes “for” anti-social behavior and metabolic conservatism. In the absence of a signal favoring the expression of these genes, they are nonetheless granted a random chance to “veto” the prevailing signal.
Combined, 17 and 18 explain why (pro- and anti-) social traits do not disappear over time, which otherwise would lead to the constant extinction of most genomes. (For more discussion of the potential roles of Onboarded Virus-derived transposable elements in psychological function, see footnotes.17
As endogenous genes are dominated by transposable elements, the question of whether they favor short-sighted competition against viral genes (via promotion of a hostile immune system) is interesting. Long-term, endogenous transposable elements would appear to benefit from promotion of the viral genome - as they themselves may still be in it, or may enter it in the future, leading to the colonization of other super-genomes. A multicellular immune system designed to facilitate the survival of endogenous and viral genes alike would seem to lead to the greatest flourishing of endogenous genes. However, an argument that promotion of viral genetic diversity is critical to the survival of all genes residing in the endogenous genome is wanted. This will be provided in the next argument.
{kind=link}
The simple, binary nature of the multicellular Transposon Cycle allows for an easy representation of the two modes:
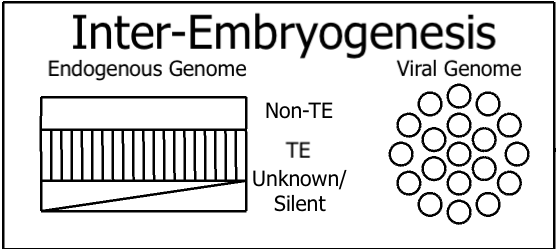
Above: Germ cell genomes, at birth, and the viral genome at birth.

Above: Germ cell genomes, after fertilization, and the evolved viral genome which nonetheless does not participate in embryogenesis.
To this, we will add Onboarded Viruses. Onboarded Viruses are stable and remain in the viral genome, but are (provisionally) the least likely to be randomly selected for access to the germ cell genome in this model. However, since they have lysogenic residence in other maternal cells, they may participate in signal planting / timing during embryogenesis, as represented by the curved line. The implication is that the selection of which onboarded viral gene contributes to signaling determines, to some extent, certain psychological characteristics in the offspring.
After embryogenesis, the offspring is naive to the circulating Onboarded Viruses, and those will be encountered early in life:

Embryogenic election is thus probably not any faster at promoting evolution than the current “errors only” model. If anything, it is a stabilizing force.
Now we will look at what happens when “stability” spells doom.
And you may find yourself living in a shotgun shack
And you may find yourself in another part of the world
And you may find yourself behind the wheel of a large automobile
And you may find yourself in a beautiful house, with a beautiful wife
And you may ask yourself, "Well, how did I get here?"19
Once in a Lifetime
One context in which a stress timer-based degradation of immunity can provide the genome a fitness advantage would be entrance of a species into new territories. In this model, the genome on the left does not experience any of the stress-signals that would suggest things are not going well in the new habitat. The genome on the right, on the other hand does. This could plausibly grant lucky offspring of the species access to advantageous local genes during transitions. The local flora and fauna is presumed to place more “edit pressure” on whatever fragment of the viral genome was transported to the new environment. Onboarded Viruses are removed, for simplicity:
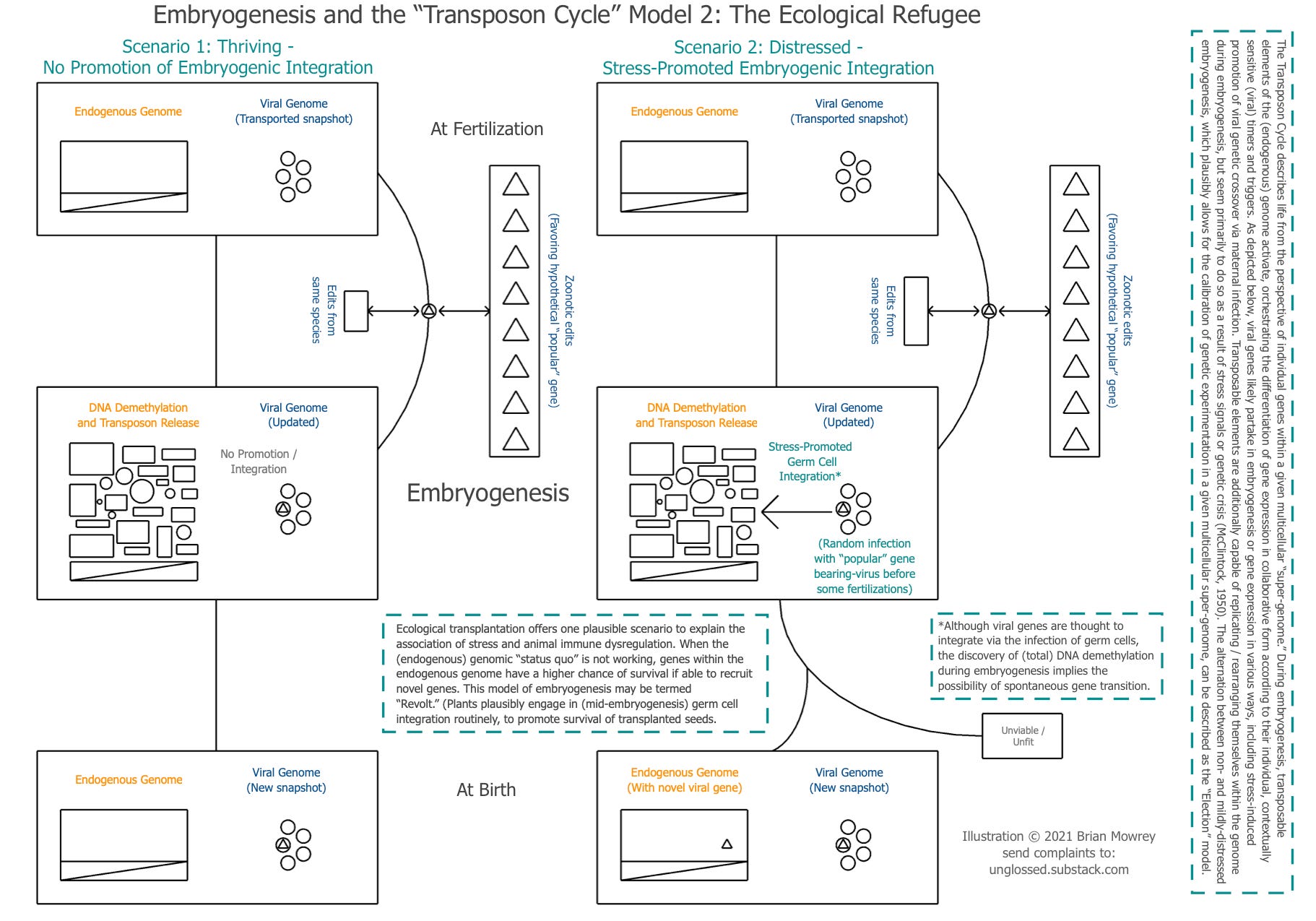
But, once again, is an “advantage” enough to overcome the self-interest of endogenous transposable elements? We probably need something more than just the challenge of a new land. To overcome the selfishness of the endogenous genome, what we need is cataclysm.
To repeat point 7 of the observational argument for Immune Equilibrium:
[T]he bacterial genome is not “invested” in the survival of the host cell. The bacterial genome is a society of genes. The regularity of cataclysmic events confronted by the bacterial genome determines that an arbitrary, exaggerated rare survival rate of 1/1,000,000 to cataclysmic events is worth an (arbitrary, exaggerated) host mortality rate of 1/10 via viral shedding, i.e. infection of bacteria by viruses not in the dormancy phase of their signal/timer pathway. In fact, only such a scheme as this can explain the successful conquest of Earth by organic matter.
The second reverse-teleological argument for Immune Equilibrium merely asserts that this is obviously true for multicellular genomes as well. There is no extant multicellular genome which has not passed through 100,000s of cataclysmic changes to environment and, subsequently, sudden reduction of fitness of the prevailing genomic order supported by embryogenic election.
Endogenous transposable elements, regardless of how “selfish” they are in temperament, cannot rely solely on the information contained within the endogenous genome to safeguard against the next cataclysm. However bleak the prospect of survival will be, survival depends on the chance being better than almost infinitesimal - again we can apply the exaggerated rate of 1/1,000,000 for the individual, or 1/2 for the species. Cataclysm implies either a situation where a genome is stranded in an environment it is not suited for, or when conditions in multiple ecosystems suddenly become so hostile that a large portion of existing plant and animal populations trend toward extinction in concert.
The most recently visible of these events in the past was 66 million years ago at the end of the Cretaceous period, when more than half of the plant and animal genomes in existence were rendered obsolete at once, but the fossil record suggests multiple iterations of even worse cataclysms before then.
Thus, like prokaryotic genomes, many multicellular genomes plausibly rely on stable, dormancy-timer equipped viruses to confer traits robust enough to ensure resilience to absence of resources, rather than risk placing those elements in the endogenous genome where they will be subject to natural selection. Plants and non-mammalian genomes arguably leverage some degree of at-will access to such traits, explaining the “common” appearance of extremophile species in those realms - those whose eggs are capable of waiting out the temporary destruction of their own biome. Plants, fungi, insects, and worms, especially, are the most plausible candidates for early multicellular rehabilitation of devastated biomes.
Thus, there is an argument for the promotion of stable viruses in the multicellular genome; and mere promotion of stable viruses plausibly guarantees the creation of less stable viruses. This places the immune system in the quandary of the first argument, meaning that staggered vulnerability is required for all non-immune-evading viruses, to avoid incentivizing the viruses to achieve immune escape, etc.
But mammals, of course, cannot rely on such tactics. The stable, resiliency-conferring traits which could apply to mammalian genomes, as listed above, seem limited to psychological anti-sociality and metabolic conservatism, not conversion into a spore that remains viable for 100 years. Yet here we still are - and in fact, we were forged in cataclysm.
Our ancestral, egg-laying family fell from 15 million years of dominance when it was struck by the greatest extinction event in historical record, the Permian–Triassic, in which 70% of land animals are thought to have died. From here, the ancestors of mammals passed through roughly 185 years of sustained pressure, including another mass extinction event at the end of the Triassic, as they headed toward the threshold of the famed event which would allegedly extinguish the dinosaurs (no, we meant “turn them into birds”). It was in this trying era that the radical departure from the egg was made. By 160 million years ago, mammals had developed fur and placental development was closer to the modern form than the marsupial pseudo-form; and at the same time, Pangea was splitting apart. By 100 years ago, all modern mammalian ancestors began to diverge into the camps which still define them, yet no fossil evidence supports that the placenta had finished developing to the modern form before 66 million years ago. And what lucky event coincided with the adoption of this genomic miracle? The extinction event 66 million years ago at the end of the Cretaceous period, when more than half of the plant and animal genomes in existence were rendered obsolete at once. Here we must ask: Did the end-Cretaceous event cause the innovation of the placenta, or did that innovation - after having been caused by nearly two-million years of repeated crisis targeted at the mammalian ancestor genome - cause the cataclysm directed back outward toward the rest of creation?
The mammalian genome had taken off the table the most time-tested tool for genomic resiliency in creation, yet it persevered through a period of general immiseration. And during all the innovation which followed - within 100 million years of abandoning the egg, mammals became unrecognizable even to their monotreme brethren - it is clear that the mammalian genome was running toward, not away, from genetic disruption. And if the competition of viral genes with endogenous genes is one source of genetic disruption, this implies that such competition if anything was not discouraged by the development of the mammalian immune system. This all implies that in some way mammals had discovered how to increase their own rate of genetic experimentation in response to stress signals while still developing ever more complex and longer routines of embryogenesis.
Even more miraculous - or, if you are a Darwinian, disturbing - is that the innovation of the placenta appears to have leaped across every extant mammalian ancestor at once, transforming them into more modern models.
These are all facts which, ever since the replacement of God with Darwinian natural selection, have caused us no end of mystification. How are there so many different types of life, with so little common ground between them? How, even among the forms with common ground, do so many features appear to have arisen out of nowhere? How, among the forms without common ground, have so many features seemingly leaped between brethren or across entire realms?
That these key leaps were shared between different genomes with viral genes, is an obvious answer. Endogenous retroviruses make up large portions of most genomes we have looked at; in fact, if all transposable elements are the progeny of ancient retroviruses, they would make up just over half the human genome. Among the phenotypes to which we owe an endogenous retrovirus, for example, is syncytin-1: The cell fusion protein which is critical to the formation of the human placenta, and which we may have just indirectly targeted for autoimmune destruction with the Covid vaccines.
The association between stress, viral infection leading to lysogeny in the germ cell, and, thereby, up-regulated levels of genomic experimentation is intuitive and supported by what glimpses of the historical record we can make out. When the mammalian genome - the society of genes - senses that “what we are doing isn’t working,” it is plausible that the orderly election model of embryogenesis is replaced by something like revolt, where all social order is reorganized, and convention is replaced by the application of ideas old and new alike.
Thus, if it can be demonstrated that embryogenesis is internally capable of promoting radically elevated levels of transposable element activity in response to stimuli, we will have proof that the society of genes is intrinsically capable of revolt. This was demonstrated by Barbara McClintock in 1944. In fact, it’s how transposable elements were discovered.
Part 2 will discuss McClintock’s experiment in more depth. Although her findings were ignored for decades, what she observed with maize seedlings was earth-shattering: By breaking off the end of a chromosome, she could prompt genes to rearrange themselves throughout the rest of the genome by apparent autonomous will, resulting in a degree of variation between related plants that had not been observed in any experiment in the history of biology to date. By sending the genome into crisis, she prompted genomic fluidity, and innovation.
If the multicellular endogenous genome has a built in sensor for crisis, it could plausibly have a built-in sensor for lifetime and immediate maternal stress. If the immune system calibrates itself to weaken under that same stress, the multicellular endogenous genome could plausibly have a mechanism for inviting innovation from the viral component. If the mammalian immune system achieved competence at promoting the proliferation and diversity of quickly-evolving viruses that shared receptor compatibility with other mammals, it further could share those rare viral contributions which were successful across multiple species at once.
Only such a stress-toggled viral innovation pathway into the multicellular embryogenic revolt state coherently explains evolutionary leaps in general, and the evolutionary history of mammals in particular. Further, although it was not delved into much here: This is also the only plausible explanation for the simultaneous development of sophisticated social neurological phenotypes amongst animals that were already hundreds of millions of years apart from their last common ancestor.21
Thus, we may offer two models for embryogenic revolt as a technique for genomes to individually and socially innovate out of cataclysm: First is the internal cataclysm - this one is already experimentally demonstrated by McClintock: in response to damage to the genome, transposable elements proliferate. Plausibly, formerly inactive genes are exploited for help/repair. Additionally, although it was not a part of her experiment, I made the supposition that nearby wild viruses are often available for potential employment. This is especially plausible for bacterial, fungal, and plant spores, for which environmental damage is both frequent, and previously shed viruses abundant
Second, the mass extinction event, when the genome that correctly discovers the viral gene that provides the embryogenic solution to a problem - represented by the viral gene “triangle” - plausibly goes on to spread that viral gene to all compatible species, with the result that multicellular endogenous genomic self-interest universally supports the most diverse and malleable Collaborative Genome possible as a hedge against cataclysm. In these models, Onboarded Viruses are removed for simplicity.


To be concluded in Part 2…
You may ask yourself, "What is that beautiful house?"
You may ask yourself, "Where does that highway go to?"
And you may ask yourself, "Am I right? Am I wrong?"
And you may say to yourself, "My God! What have I done?"

Byrne, David. Frantz, Chris. Harrison, Jerry. Weymouth, Tina. “Life During Wartime.” Fear of Music. Sire, 1979.
This is a simplification. Some distinction, or perhaps a great deal, may be argued based on dynamics of genetic competition within the super-genome. Since endogenous genes can be selfish/assertive and dependent/submissive, however, there is little “moral” distinction that can be made between the endogenous and viral element of the super-genome. Neither one is more “the true genome.” In fact, intracellular, biomolecular gate-keeping is plausibly more aggressive for endogenous genes than for viral genes.
It is possible, additionally, that antibodies to these viruses remain higher as a response to dormancy, mirroring the tendency of bacterial phages to confer immune capabilities to their hosts during lysogeny.
The extent to which dormancy-deficient viruses might coexist with bacteria, and the degree of bacterial immune competence against such viruses, is an interesting question. As resistance to “ride-on” viruses has been observed as one of the common features endowed by lysogeny, it seems that dormancy-deficient viruses would generally be kept in check, if not out-competed to extinction. I think “kept in check” is more plausible, and suspect that dormancy-deficient phages are instrumental in horizontal transfer between bacterial species.
This could also be termed the Social Genome; however, as this essay makes heavy use of the analogy of “the society of genes,” I chose the more cumbersome construction. Although I could find one paper by Domingue, B. and Belsky, D. from 2017 essentially using the term “Social Genome” in the matter I propose for Collaborative Genome, their model does not incorporate viruses or the microbiome.
See “Die Herd.” For the list of predictions etc., see Footnote 4.
Thus, all attempts to “vaccinate cancer away” are hogwash. The virus only appears because the host tissue is distressed to begin with. Here, as elsewhere, Immune Equilibrium subsumes Host Theory.
Tenet 3-1 provisionally does not apply to rare-spread pathways, namely cross-species injection-based viral infection such as Dengue virus, which seems to produce a semi-nonfunctional immune response.
See Koonin, EV. “Viruses and mobile elements as drivers of evolutionary transitions.” Philosophical Transactions of the Royal Society B: Biological Sciences. (2016, August 19.) p. 371.
For example, compare the musings on Polio offered above and expanded in footnote 12 of “Die Herd,” with the segment appearing on the September 9 episode of the Kate Dalley Radio Show. Here she alludes to a possible interplay between penicillin and the polio severe outcome surge; but I have never seen anyone else suggest such a link in my life (although I did post it in a comment online two weeks before publishing “Die Herd”). Dalley, whose program I only encountered for the first time on Friday, appears to possess a brilliant mind, one which is a year ahead of me in seriously researching the behemoth of false narratives that is The Pandemic™.
Other examples of this collective unconscious synergy is the simultaneous publication of my Theory of Covid Vaccination Immune Response in “Boostermania” with the widespread “re-discovery” of previously neglected “mucosal immunity” by the more competent and marginalized of the expert contacts of “science journalists;” the sudden critical mass emergence of awareness that Covid-vaccinated will immediately be classified as “unvaccinated” once the actual unvaccinated are successfully eliminated from society, if not before, as discussed in “Fully Vaccinated [Revokable];” and the removal of “produces immunity” from the CDC definition of “vaccine” less than a week after I pointed it out. Perhaps this is better evidence of a powerful blind-spot in my own thinking, which renders me literally incapable of seeing arguments that do not coherently match my current model of reality. Perhaps I, alone or with the rest of you, am in the midst of a Matrix meltdown preceding a reboot, where the simulation already knows in advance what I will decide next. Whatever the case the result is that the most dramatic changes to understanding of what is ostensibly a rapidly-evolving situation immediately come to appear obvious, even trite.
Ruechel, Julius. “The Snake-Oil Salesmen and the COVID-Zero Con: A Classic Bait-And-Switch for a Lifetime of Booster Shots (Immunity as a Service).” (2021, September 2.) juliusruechel.com.
Ruechel’s essay incorporates both his own independent construction of Tenet 3, reconstruction of the history of the Herd Immunity Fraud, and description of the Booster Treadmill-based Biosecurity Dystopia, the last of which I alluded to both in the sub-title of The Natural Immunity Illusion Illusion, Pt. 1 and more extensively in “Natural Immunity and the Inquisitor’s Cell” and elsewhere. I am not sure I would link the Herd Immunity Fraud to any potential intentional campaign of rendering natural immunity obsolete, however - even if such a campaign certainly exploited the pre-existing model of the Fraud. The Fraud pre-dates any possible glimpse of such an application (see “Die Herd”); whether the resurrection of the Fraud in the 90’s was partly prompted by such nefarious designs is unknowable.
Once it is (self-) evident that the genome is functionally competent at direct and indirect self-authorship, further investigation into the mechanism of these processes is unproductive: All molecular biology is linguistic and relativistic; the relationship between genes and biomolecular outputs can be adequately observed without nitpicking the physical underpinnings.
“Virus” and “theory” are two other terms which have not been applied here in the strictest sense. “Virus” is occasionally used when discussing phages, as phages are, of course, merely a human-imposed category of viruses. And this journal always employs the lay definition of theory as “any set of ideas to explain an observed phenomenon.”
Though, it was confirming the discovery of the urinary microbiome - predicted by my intuition of Immune Equilibrium in 2011 - that led to writing “The War on Pee.”
Byrne, David. “Perfect World.” Little Creatures. Sire, 1979.
All other elements of the RNA biomolecular machine can be said to have “nonrealizable” self-interest. Yet even this is wobbly. A protein is a set of RNA biomolecular machine-compatible information that would “benefit” from perpetuation through time, but it is doomed to be wielded as a physical implement to serve the self-interest of genes - yet, much like the genome of a domesticated cow, the protein’s information proliferates excellently into the future cells of its operating genome.
I.e., a “true parasitic gene.” However, since the embryogenic phase of the Transposon Cycle typically results in the perpetuation of original chromosome structure, it is plausible that the conventional view of Transposable Elements as becoming super-repeaters without any corresponding value in gene expression appears outdated.
DNA demethylation, and therefor transposable elements, are implicated in all elements of biological thinking and memory formation, including immunological discernment and memory formation and, literally, thinking and memory formation, as well as addiction and pain.
It is thus at least possible, if not incredibly likely, that the overwhelming and endlessly-remarked-upon resemblance between linguistic patterns, music, and viruses is, dizzyingly, a manifestation of the fact that thoughts are produced in the nervous system by viruses.
This would lead to an even stronger, more direct model for Onboarded Virus influence on personality, in which the Onboarded Virus genome is the endogenous psychological genome, and all immune-evading viruses dormancy-timers are designed not just to turn off once reaching the nervous system but to get there to begin with.
Either a maternally-stimulated or direct, post-birth influence of Onboarded Viruses on psychological development is the most plausible explanation for a link between vaccines and a neurological disorder characterized by an inability to perceive social patterns and master communication. I do not know if any link between vaccines and impairment of transposable elements whose behavior is mediated by or which are directly derived from the (onboarded) viral genome has been previously suggested in anti-vaxx literature.
I am not sure at the moment which model of Onboarded Virus-mediated psychological regulation I find more convincing. I have given more thought to maternally-stimulated, and the predicted role in diversification of temperament among live progeny (so that a given genome is always placing at least a few bids against the countervailing preferred traits).
Byrne, David. “Once in a Lifetime.” Remain in Light. Sire, 1980.
(index link anchor)
As mentioned in “Die Herd,” an early glimpse of this account for evolution is provided in Karin Mölling’s work. See: (2013.) “What contemporary viruses tell us about evolution: a personal view.” Archives of Virology.
Viruses helped in building genomes and are driving evolution. Viruses and bacteria belong to the human body and our environment as a well-balanced ecosystem. Only in unbalanced situations do viruses cause infectious diseases or cancer.
(index link anchor)
This strikes me as a profound insight about the nature of viruses, microbiomes, and the immune system that will ultimately be proven true.
Germ theory is an extremely reductionist paradigm; as such its utility is perhaps limited to those pathogens which have not (yet) entered into some sort of equilibrium with the human supergenome. The rabies virus comes to mind; perhaps this was also true of smallpox.
Eradication is only achievable - and should only be the goal - when the interests of the virus and the human host are sufficiently divergent that stable equilibrium is unlikely or too costly. (e.g. rabies which transmits by hijacking the brain to cause victims to bite others) In most cases the goal should instead be to facilitate the establishment of equilibrium with a minimum of suffering and death (e.g. the Swedish approach to Covid-19).
"For viruses, the immune system’s “goal” is to balance host and viral survival. This is why the immune system acts like that is its goal. Antibody levels are meant to drop after a cellular immunity rest period, as a bid for reencounter and re-spread with a given viral challenge."
That is a profound insight, and also something that should be obvious if we could let go of our conception of the immune system as the body's Department of War. Viruses can easily mutate to become more dangerous, but they will not do so if natural selection favors those that are less dangerous. Thus it is in the interest of the immune system to *allow* viruses to replicate and transmit rather than to force viruses through millions of genetic bottlenecks in which only the most attack-resistant variants survive.
Evolution of supergenomes driven by cataclysm-punctuated equilibrium. I like it.
Regarding ''herd immunity'':
https://dailysceptic.org/2021/08/06/what-the-modellers-still-dont-understand-about-herd-immunity/
My view:
Herd immunity, the widespread and temporally distributed spread of microbiological updates, has been happening for ages through precisely those people who have recently been stigmatised as asymtomatic disease spreaders. It is precisely because someone is healthy and moves freely, but nevertheless spreads viruses and bacteria to a small extent continuously through encounters and contacts, that an always up-to-date mixture of the most diverse "pathogens" can spread in the first place in an environment whose infection level is very low but sufficient to trigger T-cell actions that continuously negotiate immunities. This constant biological up-to-dateness cannot be provided by any injection, even if one were to be "blessed" with hundreds of injections every month. Immunity means being able to meet each other by permanently negotiating the capacity for closeness and the need for distance. For over a year now, we have allowed all this to happen to us unilaterally by means of distancing, and we actually imagine for the most part that this will remain without consequences. Without asymptomatic encounters, there is no healthy herd immunity, although the word no longer fits today.
Our most pressing bio-logical problem is that the mother tongue of life is influenced within a very short time by various foreign languages (mRNA-injections, artificial electromagnetic fields, ...) that our bodies are not given enough time and context to understand what is going on. Therefore more and more viral intervention is on its way to fit our (mis)understand of life into what life really is. Thus more and more symptoms. Thus more and more inventions to counter those symptoms. Thus a vicious circle of our own making goes on and on. Blind and deaf we create what we want to avoid.