“Imprinting” was demonstrated in breakthrough-infections with BA.1/2/5.
Convergent evolution is observable in circulating BA.2 and BA.5 variants.
“Imprinting” is driving this convergent evolution.
The result will be that, after a few more mutations, BA.2 and BA.5 variants will achieve total escape from neutralization by the breakthrough-infected.
While “antibody evasion prediction studies” are generally a snooze-fest, the outlandish claims regarding immune-mediated evolution deserve inspection. And so, let’s go.
The set-up
Blood draws were taken from triple-inactivated-Covid-vaccine-dosed patients either without infection or “hospitalized” (quarantined, presumably) for infection with an Omicron type:
Post 3 Inactivated Covid vaccines, adults predominately aged 20s-50s:
Uninfected: 40 subjects, sampled about 4 weeks after 3rd dose2
BA.1-infected: 50 subjects, sampled ~40+ days after admission
BA.2-infected: 39 subjects, sampled 38 - 77 days after admission
BA.5-infected: 23 subjects, sampled 16 - 36 days after admission
Also:
A non-vaccinated, infected set of samples came from somewhere.
As we can see, there was variation in the time between infection and sample-collection, with good time-periods for new immune responses given in the BA.1 and BA.2 set and less ideal time-periods in the BA.5 set.
Next, “a lot of various kinds of tests” were then done.3
Plasma was tested for neutralization of pseudoviruses with a gamut of trendy BA.2 and BA.5 mutations and cool names (XBB, BQ.1.1, etc.)
B Cells that Wuhan or variant Receptor Binding Domains attached to were isolated to make a monoclonal anti-RBD antibody “library” for each patient. (This is a common approach in “imprinting” studies.)
From here, there were inferences, inferences, and more inferences.
Part 1: Imprinting
Context: “Imprinting” and the Covid vaccines
A review of terms and prior evidence for “imprinting” with the Pfizer/BioNTech Covid vaccines is in the footnotes.4
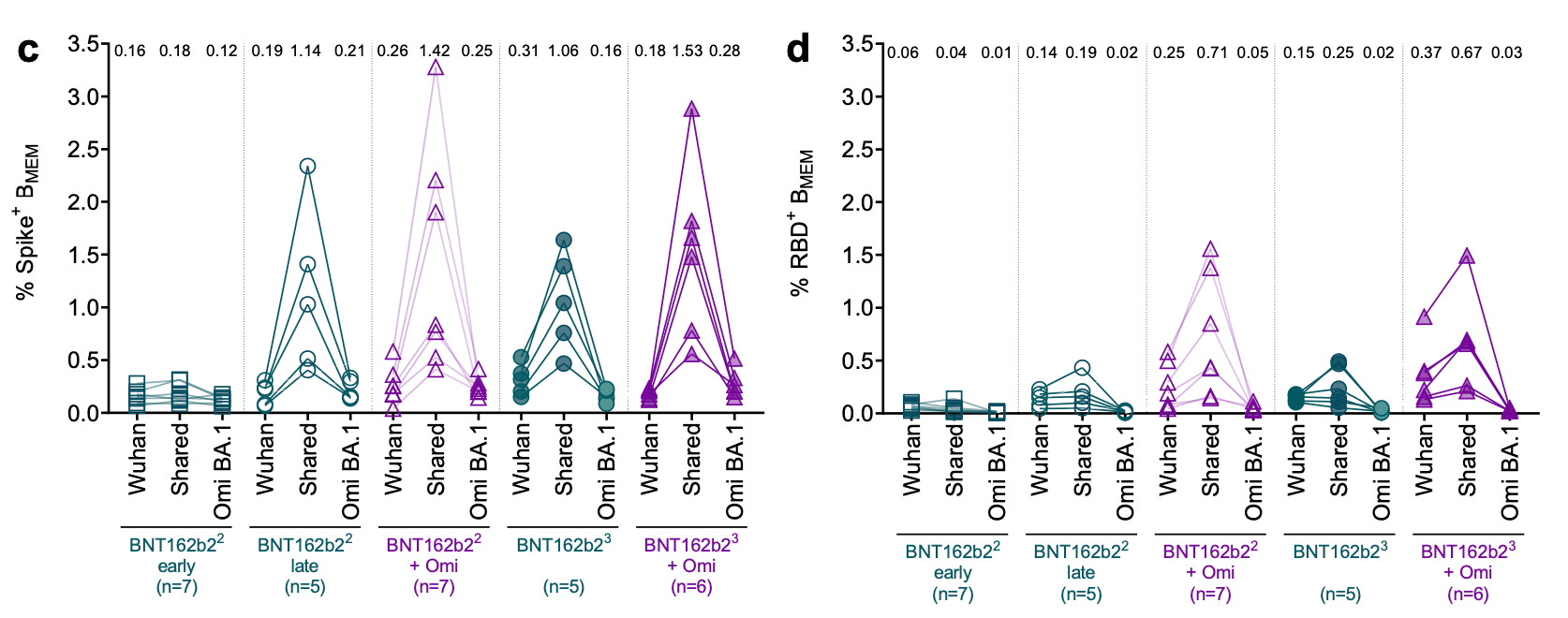
Results: “Imprinting” antibodies are not universal after inactivated vaccines.
Unlike with the weird, experimental Pfizer/BioNTech mRNA vaccine, the inactivated CoronaVac5 vaccines do not seem to leave recipients stuck with their original “team” of super-cross-reactive memory B Cells.
Cao, et al. Fig 3. In (d), B Cells are sequenced to produce monoclonal antibodies, resulting in a lower percentage of variant-specific results since clonal duplicates are not counted. BA.5 convalescents were sampled more recently after infection, and so have higher overall B Cells but fewer models of specific and cross-reactive antibodies (or, higher clonal expansion is still reducing the representation of rarer clones).
This, as reviewed in the Context footnote, is in stark contrast to results after BA.1 infection for the Pfizer/BioNTech-injected, where almost no BA.1 Receptor Binding Domain binders were “specific” (did not also bind Wuhan RBD).
In (d), we see that when these non-cross-reactive, variant-specific B Cells are examined for how many times they have edited (mutated) the genes responsible for modeling their B Cell Receptor and corresponding antibody, fewer edits (SHM’s) are found than for the cross-reacting B Cells. This means these B Cells are new recruits just out of the naive B Cell pool; they are a truly novel immune response to the variants that infected these vaccinated donors.
Naturally, besides the new, specific B Cells there has been an expansion in Wuhan-binding B Cells that also bind to the variants. This is how immunity is supposed to work; protective prior responses are used, not thrown out the door like last year’s iPhone. Notably, the vaccinated have much higher overall B Cells that can bind their variants than the “BA.2 infection without vaccination” group (more blue dots in the diagonal oval)6; this might not mean anything in terms of protection against actual re-infection but it clearly fails to show a detriment from imprinting.
Of course, the authors don’t see it this way; to them, this is a big win for “OAS.” After all, OAS is true no matter what, and so this must prove OAS is true (emphasis added):
Similar to that reported in BA.1 breakthrough infection, immune imprinting, or so-called “original antigenic sin”, is also observed in BA.2 and BA.5 breakthrough infection. Post- vaccination infection with BA.2 and BA.5 mainly recalls cross-reactive memory B cells elicited by wildtype-based vaccine, but rarely produces BA.2/BA.5 specific B cells, similar to BA.1 breakthrough infection
30% is now “rarely”?!
Whatever.
Conclusion 1:
There was some “imprinting,” which means there wasn’t really “imprinting” (see terms discussion in footnotes).
Antibodies Narrower?
The authors claim that their results show that this partial vaccine “imprinting” results in a narrower B Cell / antibody response to the respective Omicron variant. I would note the comments included in my caption above: A lot of the “narrower” breadth in the BA.5 group seems driven by sample timing. It is thus important to see how the authors measured “narrowness.” They provide this figure:
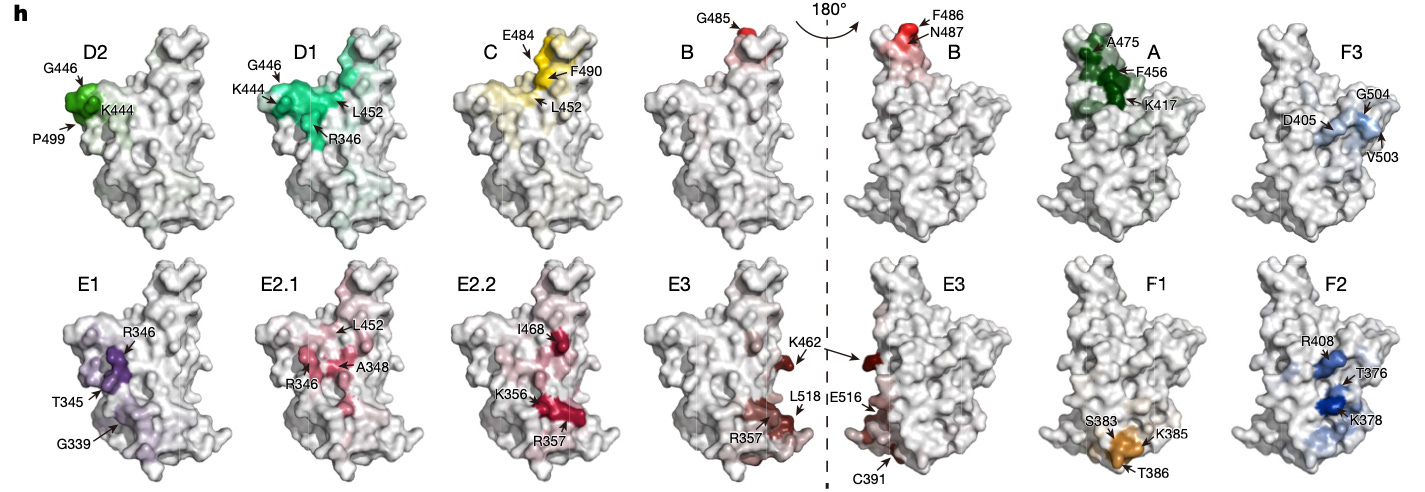
We can essentially imagine (e) as a map of the receptor binding domain’s surface as seen by an antibody, the same way the Earth’s surface could be peeled to make a 2D map; except the map also represents alternate, "remodeled" Earths (the Omicron variants7).
Compared to “WT convalescents/vaccinees,” the breakthrough convalescent antibodies heavily focus on the southern and western regions, which per the results in (g) are antibodies that do not compete with the RBD for ACE-2 binding and are not “neutralizing” (do not protect cell cultures from the virus). This, notably, reflects the fact that the more ACE-2 interacting, neutralize-y F, A, B, and C regions are highly mutated in the Omicron siblings (and D1 in BA.5); while D2 and E2/3 regions are more conserved. With the exception of the expansion of C-region antibodies after BA.2 infection, it seems that there has been a predictable reinforcement of cross-reactive antibodies in conserved epitopes, with a decrease in the breadth of truly neutralizing antibodies.
Sample raw data from Supplementary table 2. Post-infection antibodies found to bind to the BA.5 Receptor Binding Domain are cloned and evaluated for neutralization against D614G (blue), BA.2 (red), and BA.5 (yellow), lower numbers neutralize better.
Conclusion 2:
Breakthrough infection did not result in as many clone “models” of B Cells / antibodies that bind to the top regions of the variant Receptor Binding Domain. However, since just one neutralizing clone can pump out functionally any quantity of antibodies, this has no intrinsic relevance for immunity — accordingly, there was no evident effect on whole plasma post-infection neutralization of the corresponding variant.8Also, the disparate results for BA.2 and BA.5 suggest that a later time-point for the latter will reveal greater antibody diversity in the RBD (a point that has been relevant for all the “imprinting” studies reviewed this year).
At which point we could say, so what?One neutralizing antibody is enough to do the job; how many (if any) does a person need?
As if specifically in order to provide an answer, the authors turn to convergent evolution.9
A final note before moving on, is that comparison with “WT” responses is not necessarily an illuminating way to “quantify” the antibody response to the coveted F, A, B, C regions of the Receptor Binding Domain. Different viruses may prompt different immune responses, especially two variants with such dissimilar virulence. How does the number of binding antibody models for these regions compare to the unvaccinated, BA.2 infected group shown in Fig 3c?
If you bet five million dollars on “the authors do not include those results,” today is your lucky day. See the nearest Walmart manager to collect; they will know I sent you.
Part 2: Like a Convergion
The authors begin the paper by observing, hyperbolically, that BA.2, 2.75 and 5 have recently picked up similar mutations (emphasis added).
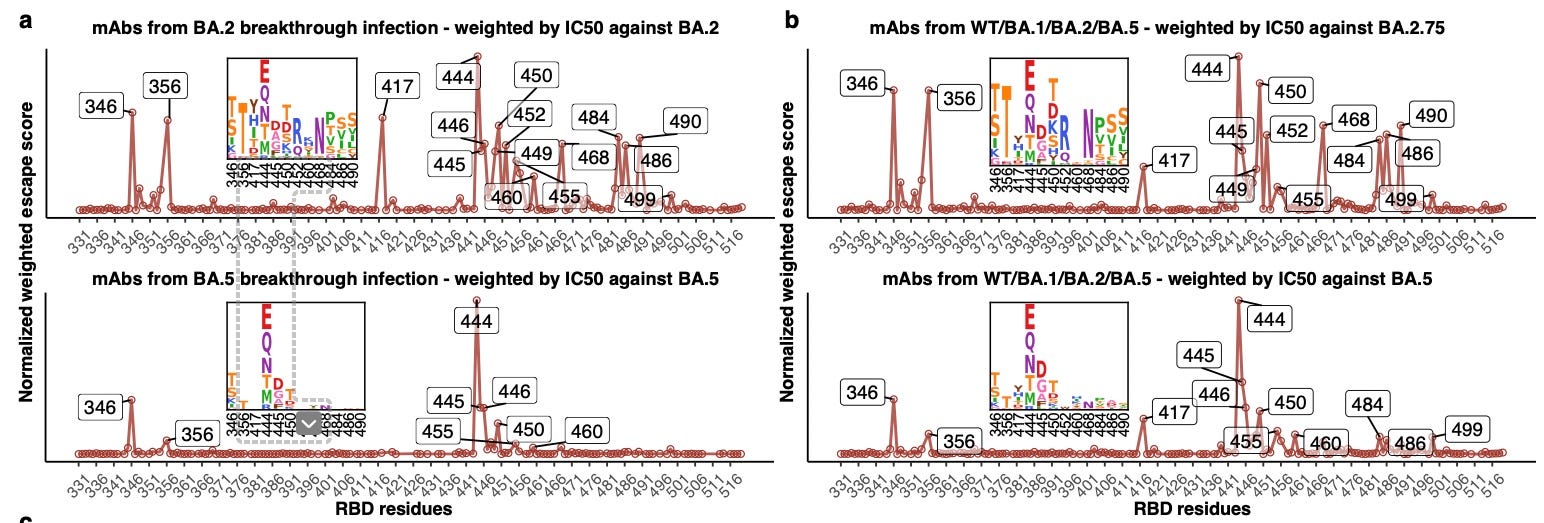
Continuous evolution of Omicron has led to numerous subvariants that exhibit growth advantage over BA.5. Such rapid and simultaneous emergence of variants with enormous advantages is unprecedented. Despite their rapidly divergent evolutionary courses, mutations on their receptor-binding domain (RBD) converge on several hotspots, including R346, K356, K444, L452, N460K and F486. The driving force and destination of such convergent evolution and its impact on humoral immunity established by vaccination and infection remain unclear.
This is, obviously, idiotic. The SARS-CoV-2 Receptor Binding Domain has featured “hot spots” from the start. This plot could have been created this same time two years ago featuring residues 341, 435, 458, etc.10 These all proved worthless for prediction upon the simultaneous emergence of the "Variants of Concern" at the end of the year; and who would be the least bit surprised if the same thing happens again in another month? The only difference in October, 2022, is that we are being looser with the special secret-agent code-names.
Observe that the greek letter variants featured convergent evolution, and yet this was hardly sensationalized two years ago during their simultaneous emergence. A slide from a CDC training module for variant-tracking with nextstrain and other tools calmly points out the appearance of the N501Y mutation in 3 otherwise unrelated B.1 offshoots:
https://www.cdc.gov/amd/pdf/slidesets/toolkitmodule_1.4-508c.pdf (annotated). Note that the branches for these three variants are all fictions — they do not correspond to sequence-observed iterative mutations, but like with Omicron the next year merely attempt to model a natural creation of what were more likely lab-made variants built from the B.1 backbone. More discussion in Omicron Origins.
Where was the op-y handwringing about “convergent evolution!” when it was observed in what were plausibly actual lab-created variants?
More to the point, it is not surprising that BA.2 and BA.5 are making the same mutation “moves,” given that they are both working with a similar deck and both playing on the same board. Whether BA.2 got a head-start in the post-BA.1 immune landscape by luck or due to some advantage, it quickly hit its own wall of immunity, conferring a fitness advantage to the similar, but some-moves-different BA.4/5 (same spike protein on both). These two were sequenced early this year at the same ground zero as the other Omicron siblings; suggesting they were already present in the original release.11 Once the immune landscape shifted in their favor, they displaced BA.2.
But at a certain point, around this summer depending on region, both BA.2 and BA.5 would essentially be facing the same wall of immunity and need to fish for whatever available “moves” neither of them had yet played. Essentially, they might as well be the same virus.
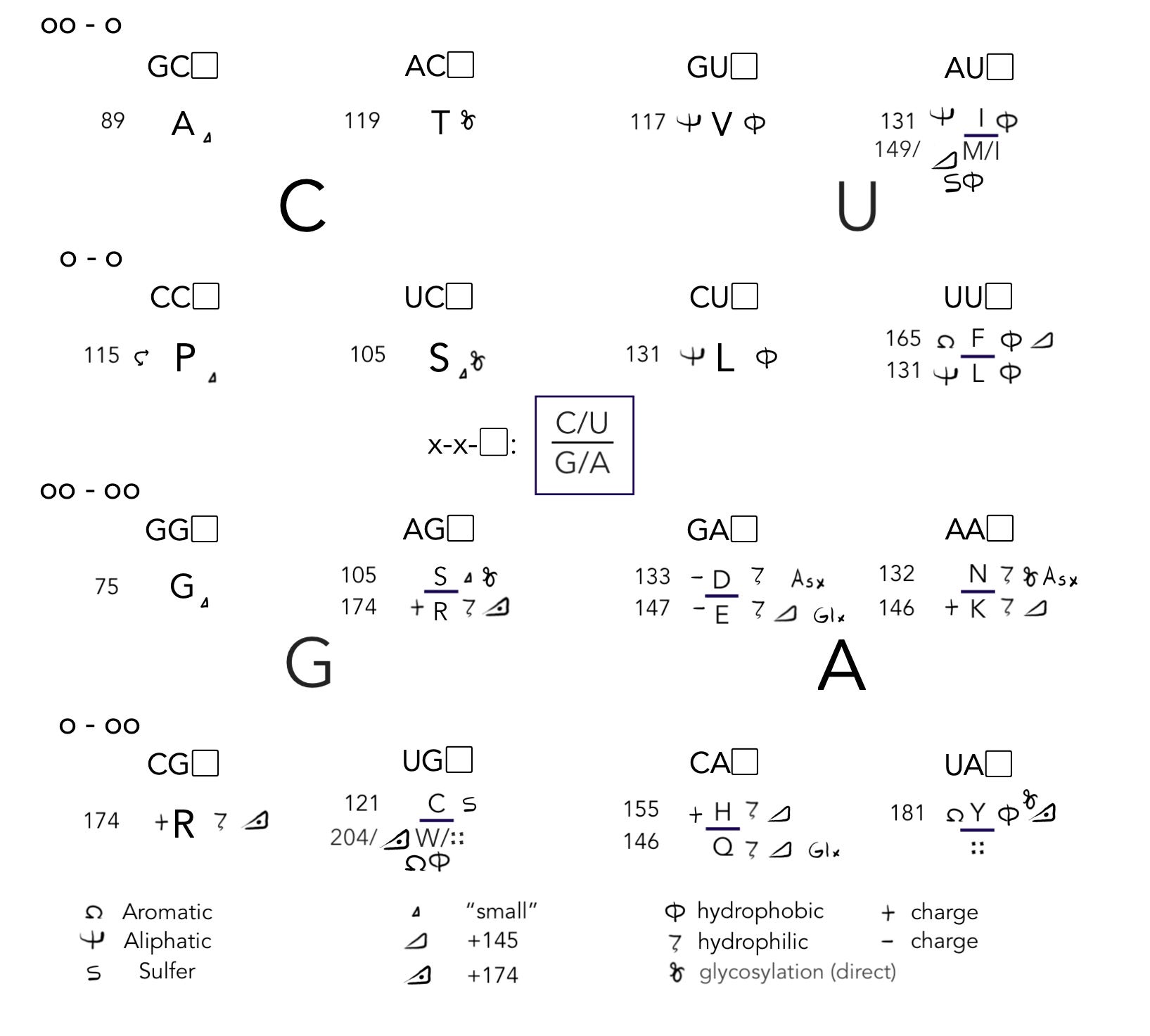
Speaking of evolution as moves, here, again, I will give the screen to my crackpot genetic code bit-map, “CUGA:”
A mutation to a single nucleotide (C/U/G/A “letter”) either leaves residues as-is or moves them to a compatible place on the board. Any given K on Earth at this exact moment, for example, can stay the same, swap to N, hop to E/Q, or leap to T/(M/I)/R in a single move. It can’t become any of the other residues without two separate moves. It is also constrained between M and I depending on whether it is actually AAG or AAA; we can think of it as having a 50/50 chance on that move.
These set-in-the-code move possibilities are the foundation of “Deep Mutational Scanning,” which lets computers game out which single mutations will have the biggest impact on a given protein’s performance. Cao, et al. (the authors of today’s paper), use this very technique to determine which mutations, if acquired by BA.2 and BA.5 would confer escape from their collection of antibody libraries.
The computer thinks that both BA.2 and BA.5, if playing against the antibody library “decks” collected by these researchers, should play 346 and 444 (as well as many others moves for BA.2, but we’ll get back to that). Sure enough, these are among the two hotspots for convergent BA.2/5 mutation!
But, so what.
The authors propose that because they have shown that 1) Breakthrough infection response reveals “imprinting,” and 2) BA.2/5 seem to have few ideal Receptor Binding Domain mutations that achieve immune escape, then 3) “Imprinting” is driving immune escape.
Huh? This is just a list of ideas, not a demonstration of cause and effect.
Convergent evolution suggests pressure, not permissiveness
How do the authors actually adjudicate whether “imprinting” is driving the limitation of immune escape moves? Wouldn’t a narrow immune response increase available immune escape moves? Especially if that narrow-ness was concentrated on the Receptor Binding Domain? For example, if post-breakthrough neutralization of the BA.5 RBD depended on repurposed, not-strongly-binding Wuhan-vaccine antibodies, then the virus ought to be able to profit from a change just about anywhere. There’s obviously implications and considerations that the authors have failed to delve into.12
What makes BA.2 and BA.5 different from each other?
Further, if “imprinting” is the story here, why would BA.2 and BA.5 have different degrees of move freedom? Without even understanding what is going on, BA.2 seems sufficient to falsify the proposed mechanism for convergent evolution. The authors show which moves it thinks D614G (representing the “WT”/Wuhan RBD) should make to escape “WT infection”-generated antibodies: It looks exactly the same as BA.2, if not slightly more limited.
Cao, et al. Extended Data Fig 8. It is not clear if “from BA.1” refers to natural or breakthrough infection.
Where’s the control group?
What’s more, any claim that “imprinting” is determining the breadth of RBD response to begin with requires comparison with a non-imprinted control: The unvaccinated, BA.2-infected. If the authors can’t or won’t show that these have more antibodies in the F, A, B, C regions than the breakthrough-infected, they haven’t proved their point.
Why does convergent evolution start before “imprinting”?!
Lastly, exactly as mentioned above, there is nothing “new” about SARS-CoV-2 converging on hot-spots. Absurdly, the authors even affirm that this already happens in the background of first-exposure immunity, by running their program on the Wuhan model of the virus. Exactly as shown in Extended Data Fig 8 above, ideal moves were just as limited; and the computer was just as predictive of real-life:
Moreover, we wonder if the real-world evolutionary trends of SARS-CoV-2 RBD could be rationalized and even predicted by aggregating this large DMS dataset containing mAbs from various immune histories. Using the mAbs elicited from WT vaccinees or convalescents weighted by IC50 against the D614G strain, we identified mutation hotspots including K417N/T, K444-G446, N450, L452R, and especially E484K (Extended Data Fig. 8a). Most of these residues were mutated in previous VOCs, such as K417N/E484K in Beta, K417T/E484K in Gamma, L452R in Delta, and G446S/E484A in Omicron BA.1, confirming our estimation and inference
The authors have thus neatly refuted their entire conclusion about “imprinting.” If immunity was already driving convergent evolution before “imprinting,” then the appearance of convergent evolution in BA.2 and 5 does not prove “imprinting” is driving the effect.
Conclusion 3:
The authors have woven an incredibly weak, flawed theory out of thin air — likely in order to drum up downloads and citations with a sexy headline. There is nothing surprising about convergent evolution and nothing in their data that supports an influence on the same from “imprinting.”
An additional post may or may not address the predictions this paper makes about future Total Immune Escape, just a few mutations around the corner (like it has been since, well, forever). For now, they fall back into the category “another day, another antibody evasion prediction study.”
If you derived value from this post, please drop a few coins in your fact-barista’s tip jar.
“Original antigenic sin” is a pair of claims about antibodies. One of these claims can potentially be called “imprinting.” It depends on who is using the i-word at a given moment.
“Imprinting”
First-exposure antibodies determine the character of future variant exposure antibodies. That is, all future antibodies generated to variants of this virus will cross-react with the first-exposure virus.
or
Some other type of imagined phenomenon that depends on who is using the term at any given moment.
OAS refers to a universal doctrine. OAS not only asserts that the first condition described above happens but that virtually no non-imprint variant antibody responses are possible in human biology.* And, a writer could use "imprinting" to refer to this alleged biological "doctrine."
“Imprinting” (doctrinaire definition) example
Bob is a human who encountered H1N1 flu one time. Thus, he can never make an antibody to a variant of H1N1 that does not cross-react with his originally encountered virus. Because of the spooky voodoo of OAS imprinting.
The doctrine is not and never has been valid. And yet if we accept that hundreds of exceptions to “imprinting” have been demonstrated in the OAS literature itself, then we can still call any case where it is true “imprinting.” Rather than being the rule; it is the exception. "Imprinting" can thus be divorced from OAS and still have a coherent meaning.
“Imprinting” (non-doctrinaire definition) example
Bob got the Pfizer/BioNTech mRNA transfection for Wuhan spike protein and now he can't seem to make Omicron antibodies that don't cross-react with Wuhan spike protein; this appears to be a case of imprinting.
Alternately, a writer might use “imprinting” to simply refer to any quantity of antibodies or memory B Cells that are presumed to be from the original virus exposure. And if the writer has a non-canonical definition of OAS, she might assert this definition as being compatible with OAS.
“Imprinting” (post-truth definition) example
Bob encountered a flu variant. Greater than 0 of his post-encounter antibodies cross-react with the first flu he encountered. Without stopping to reflect whether this even relates to the order in which his infections occurred, we can be 100% sure these “imprinting” antibodies prove that OAS is real. Pick me, teacher.
Both words are prone to infidelity. Authors often even use “imprinting” without reference to specificity; in other words the word could mean truly anything at all.
And so a writer could finally claim, "OAS and imprinting are the same." And yet, as it wouldn't actually be clear to anyone what he meant by "imprinting" or “OAS,” the statement would not be meaningful; and would reflect likely ignorance about both terms (this describes most published literature).
Second, “Imprinting” vs. “Specific” antibodies
In vintage-biology terms, "imprinting" means that antibodies to variant viruses will stick to (be absorbed by) the originally encountered virus. In modern-biology terms, "imprinting" means that antibodies to variant viruses are generated by reshuffled, re-modeled B Cells from the memory B Cell pool created by exposure to the original virus. Neither of these is a bad thing; in fact it leads to quicker and stronger (higher avidity) variant responses.** But since these variant-binding antibodies also cross-react with the originally encountered virus, they are not considered “specific.”
That’s all “specific” means — “not cross-reactive.” It doesn’t imply better performance against the variant virus; and so there is no prima facie performance penalty for not having “specific” antibodies.
The only reason anyone cares about “specificity” is because there is this stupid myth of a biological doctrine that specificity after variant virus exposure is impossible (this doctrine is called OAS).
Covid vaccine “imprinting”:
The Pfizer/BioNTech vaccine seems to cause near-universal imprinting:
So far, even with long time-periods between Omicron sibling infections and samples, all post-infection Omicron-binding B Cells seem to descend from the pool elicited by the Pfizer/BioNTech Covid vaccine, at least in a study overseen by BioNTech’s Uğur Şahin himself. See “The Actual "Imprinting" Study.”
Quandt, J. et al. Fig. 4. Samples for 2-dosed + BA.1 taken 46 days post-infection; for 3-dosed + BA.1 taken 44 days. Further discussion in previous post.
This is unlike the case in flu where the evidence shows that imprinting is, in fact, far from a universal “doctrine.” And so we have some mRNA-transfected Bobs out there.
This doesn’t mean these B Cells and their corresponding antibodies are bad at “binding Omicron;” but it still has implications for the character of the mRNA vaccines and questions about tolerance and negative efficacy (as discussed in the post).
*Francis vaguely uses “largely determined” as the benchmark for the claim made by OAS. However, since a substantial portion of antibody cross-reaction is driven by intrinsic resemblance between variants, anything other than near-universal absorption of variant antibodies by original strains would be uninteresting. For example, if having “only” 25% “specific” antibodies to a variant was enough to meet the benchmark of largely determines, then OAS could be refuted by “only” 25% of the original antibodies being “specific” to the original virus (even in advance of variant vaccination; exactly the week-41 baseline Wuhan-specificity of the macaques in that ridiculous Moderna booster study). It is thus necessary that almost zero variant antibodies are specific in order for “largely determines” to have any meaning, as in Francis and co’s idealized ferret results (see “Flu: The Lost Years.”)
**Higher-than-first-exposure avidity to flu variant vaccines was even used by de St. Groth and Webster as supposed evidence of imprinting, in 1966, as noted in my OAS Lit. Review Timeline (table).
Timing of samples is important here. For example, the BA.5 samples show more binding B Cells because they were taken more recently after infection. This also suggests that for the BA.2 group, the BA.2-specific B Cells have not diminished as a proportion of the overall anti-SARS-CoV-2 B Cell response in the medium term; but true longitudinal samples would be required to verify this suggestion.
Here the text is unclear. The caption on Fig. 3 describes (e) as representing “WT” (Wuhan) spike; the text does as well, but refers to Supplementary Table 2 which includes antibodies from breakthrough infections. Comparison with Fig 3c in an earlier paper in the series, however, makes it clear that post-Wuhan antibodies are remodeling the map.
This refers to the plasma neutralization panels that I did not review in the main text of this post. As can be seen, post-infection neutralization against variants is as high as pre-infection neutralization against D614G (Wuhan).
Cao, et al. Fig 2. That BA.5 infection so strongly immunizes against the related BA.2, but not vice-versa, likely hinges on the extra escape advantage conferred by BA.5’s F486V mutation (Kurhadea, C. et al.). Donors do not yet appear to have generated many antibodies to this mutation given the low performance against the BA.2 F486V mutant; a later sampling time-point may show stronger results. Notably, there was just one outlier with a 74-day admission-sampling interval (all others were 36 days or less). Could this be the same outlier in the BA.5 neutralization graph?
As always, the back-boost of Wuhan / D614G can not be taken as meaningful if no comparison to an unvaccinated, variant-infected response is provided.
Naturally, the authors present their logic as first having observed convergent evolution, and then embarking on a quest to decipher what is driving it. This is nonsense, considering that this paper is merely another iteration in a series that was just waiting for a sexy headline.
Why, for example, does DMS decline to recommend a change to BA.5’s 417N, even though it is the same as on BA.2? The obvious answer would be that the BA.5 convalescent antibody library for some reason is not escaped by mutations to this residue, whereas the BA.2 convalescent antibody is. However, this doesn’t make much sense; there is no difference between the two variants in this region (the “A” antigenic site). The most likely answer is that the algorithm loses resolution with a smaller set of inputs (as with the earlier-collected, smaller BA.5 mAb library), likely to avoid false positives.
This is Dr Been’s simpler take on this issue. I guess you are dissecting it further. I can detect your sarcasm at some points, but because this is all new learning for me I can’t appreciate what you’re implying fully and most of these diagrams need a longer explanation!
From other reading I gather the T cell responses and NK/ dendritic cells are more important for our surviving the virus, so does the extent to which antibodies vary really matter on a practical level, other than to produce pretty papers?
I’ve also just watched Dr Been’s video about the Wuhan body/Omicron spike manufactured virus and that implies that something in the Wuhan body is what causes severe disease. It’s a while since I read about the origins of Omicron, but (apart from it having suspiciously lots of non synonymous mutations ) I thought most of its mutations were in the spike relative to the rest of the virus?
No wonder they laugh at us. While we're looking at charts and tables, they're spraying us with something all over the world. Probably setting us up for weather catastrophe while everyone plays tiddlywinks.






. However, the p value above lets you know that there is actually a difference.")








https://youtu.be/MAnvEO4Ixjk
This is Dr Been’s simpler take on this issue. I guess you are dissecting it further. I can detect your sarcasm at some points, but because this is all new learning for me I can’t appreciate what you’re implying fully and most of these diagrams need a longer explanation!
From other reading I gather the T cell responses and NK/ dendritic cells are more important for our surviving the virus, so does the extent to which antibodies vary really matter on a practical level, other than to produce pretty papers?
I’ve also just watched Dr Been’s video about the Wuhan body/Omicron spike manufactured virus and that implies that something in the Wuhan body is what causes severe disease. It’s a while since I read about the origins of Omicron, but (apart from it having suspiciously lots of non synonymous mutations ) I thought most of its mutations were in the spike relative to the rest of the virus?
No wonder they laugh at us. While we're looking at charts and tables, they're spraying us with something all over the world. Probably setting us up for weather catastrophe while everyone plays tiddlywinks.