
The Hep B vax HIV Origin theory, pt 1
Some lackluster comments

*edit, October 3: Commenter TCC alerted me a very helpful recent book on HIV origins, namely Jacques Pépin’s The Origins of AIDS, which led to the identification of some errors in my reconstruction of pre-1980s spread — namely, I had not encountered the overt description of evidence of a lack of spread outside the Congo region before 1980. This post has since been corrected to change references to the virus’s evolution in Africa to “in the Congo region;” I have also corrected the lay description of the type of chimpanzee from which HIV derives.

Friday, renegade neuroscientist Kevin McCairn featured a startling and intriguing theory on the origins of HIV. It is not entirely novel — there is at least one older interview which I had not previously viewed. But I had not encountered it before; nor, apparently, had his present interviewee until a few days ago. Said interviewee is Aaron Baalbergen, who had already been pursuing links between vaccine research and HIV in general before encountering details regarding the first Hilleman hepatitis B vaccine (FHHBV). As said, this oversteps some previously existing investigation into the topic — to wit, Friday’s interview also featured an independent researcher who simply goes by “Nick” and has previously spoken on the subject.
The reader can find Friday’s interview of both Ballbergen and Nick by McCairn here (rumble.com), but in truth, not much substance managed to find its way into the discussion. An older and perhaps more expansive McCairn interview with Nick is here (I have yet to watch it, but am including it to avoid any misattribution of research). More usefully, the FHHBV theory is succinctly and slickly expounded in a video by Ballbergen uploaded to his twitter and to youtube. Below the embed, I will summarize:
Summary of the FHHBV theory:
If taking as plausible the default narrative that HIV crossed over to humans from apes in the 20th century shortly before becoming a recognized disease agent,
Then the obscure medical fad of cocirculating the blood of hepatic coma patients with chimpanzees, which took place in the 1970s, is obviously the most likely cause of the crossover.
The FHHBV, for related and unrelated reasons, deserves scrutiny for the outbreak in the American gay community, in African countries which continued to use it after the West switched to the recombinant vaccine (i.e.e the second Hilleman Hepatitis B vaccine), and briefly in China.
The “related” reason is that initial trial materials for the FHHBV (i.e. Hepititis-B-infected plasma) may have been prepared via chimpanzee inoculation.
The “unrelated” reason is that the market version of the FHHBV was known to be prepared using (Hepatitis-B-infected, but treated) human plasma.
In fact the potential link between the FHHBV (used heavily and almost exclusively among American gays) and the HIV epidemic is a historical open secret — the concern was raised from the earliest cases, and was the de facto motivation for discontinuing use of the FHHBV in everywhere but Africa.
In this post, I will discuss why this theory depends on some probably incorrect ideas about how recently HIV began spreading in humans. In a second post, I will discuss the interesting idea that the FHHBV nonetheless was a vector for the widespread dissemination of the virus that led to the AIDS epidemic in gays et al.
Shorting the chimpanzee transfusion angle: HIV is probably ancient
Now I will offer what the subtitle promised would be lackluster comments on the notion that HIV must have resulted from admixture of blood with chimpanzees. Ballbergen is correct that if HIV crossed over from apes in the 20th Century, medical experimentation that openly mixed chimpanzee blood into humans in the West is more likely to have been the vector than some wild adventure in the African jungle.
But of course, this simply runs into and raises to light the lack of coherence, plausibility, and sophistication in the default narrative of the (recent) zoonotic origin theory of HIV.
The (recent) zoonotic origin theory proceeds from the apparent novelty of the disease as it was experienced among gay men after the American sexual revolution. Rather than behold the obvious — new host social patterns of sexual behavior and drug use were producing new1 manifestations of a previously unnoticed virus — scientists and the media pursued a narrative in which a new human virus must have appeared out of nowhere, and surely anyone infected with it would experience the same disease as being observed amongst Western gays and various drug using communities.
The notion that HIV was a new human virus could be easily if unwisely entertained in the 80s and 90s when sequencing was still a new and limited art, but is obviously provincial and implausible in light of broader knowledge which has existed now for decades without widespread recognition.
Too diverse
HIV is first of all too genetically diverse to have plausibly originated in a recent cross-over from any related animal virus (i.e. Western-chimpanzee and sooty mangabey SIV, in the cases of HIV-1 and 2, respectively). By “recent,” I refer to any timeline that equates the earliest recognized infections with the time of cross-over — really, anything after 1950. Regarding the use of the “SIV” term, I do not wish to appear to be commenting on any relationship between the pathology of the disease in other primates vs. what we call AIDS in humans. SIV is here merely meant to denote a retrovirus platform which thrives in primates, and which phylogenetic analysis finds to be the ancestor of human HIV. The topic of how SIV manifests in other primates — its pathology — is not being raised.
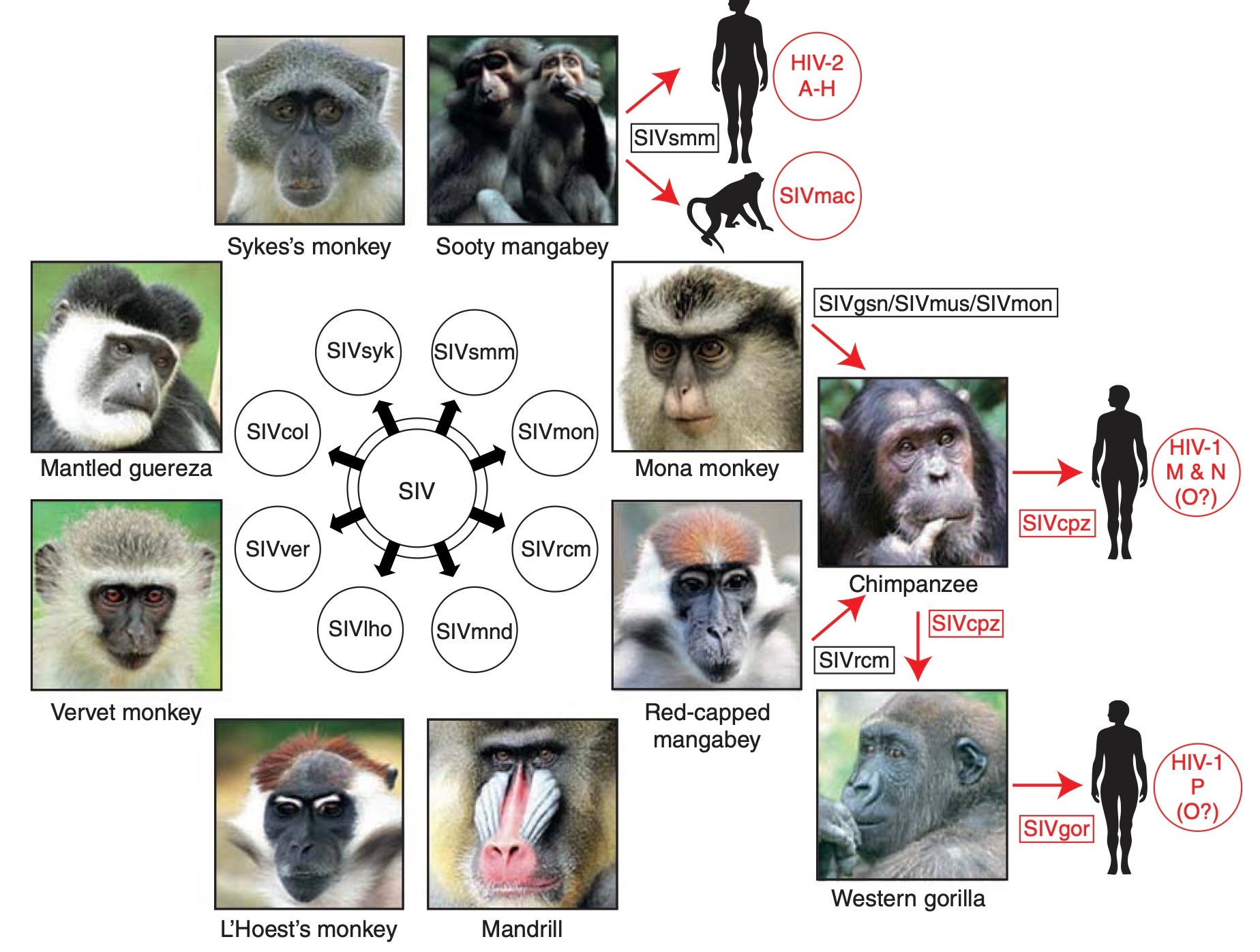
It is true that the full genetic diversity of HIV was at first limited to Africa — specifically and importantly, to the Congo region — but even this still precludes the possibility of cross-over into humans after 1950. This is true in terms of there existing multiple groups and in terms of the diversity in the M group, which is the type which predominately has spread beyond Africa and which is more pathogenic (due to surpassing a specific evolutionary barrier in human tetherin).
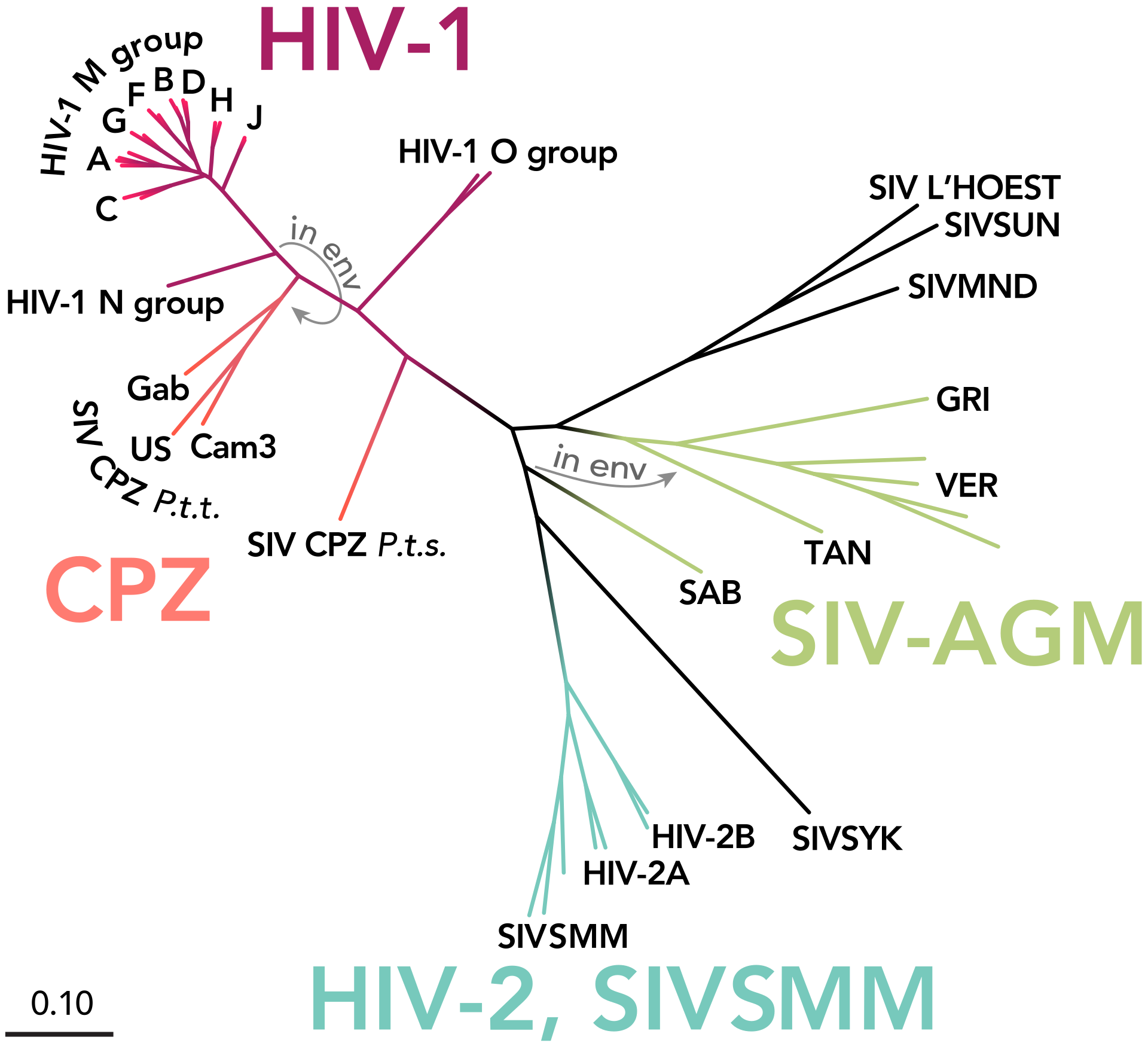
HIV-1 M specifically has eight to ten subgroups which seem to have all been in place before anyone began isolating or sequencing the virus. An early indication of this comes from two samples preserved from Kinshasa in 1959 and 60, one of which falls in the A2 subgroup and the other which falls into D. These two samples have been the subject of multiple reevaluations of the time of HIV-1M’s introduction into man. Michael Worobey et al., in a 2008 paper, write:
Our estimation of divergence times, with an evolutionary timescale spanning several decades, together with the extensive genetic distance between DRC60 and ZR59 indicate that these viruses evolved from a common ancestor circulating in the African population near the beginning of the twentieth century; TMRCA dates later than the 1930s are strongly rejected by our statistical analyses.2
Granted, these same papers were to some extent a reply to the accusation by multiple authors in the lay press that polio vaccines and associated trials potentially originated the cross-over of HIV-1 from chimpanzees to man; therefore the reader may infer some measure of conflict of interest on the part of the authors. However, there is nothing really to be argued against their points beyond the incorrect early understanding of the virus that largely persists in the “default narrative” today — i.e., that it wasn’t extremely diverse from the start.
Why diverse in humans?
Before I move on to arguing that Worobey, et al.’s estimate is perhaps too conservative, we must tackle the question of how we can be confident that all the pre-existing genetic diversity in HIV-1M developed in humans rather than in Western chimpanzees.
It is first worth re-emphasizing that the misunderstanding that HIV (and HIV-1M) was novel circa 1980 is understandable given that its diversity, at first, was limited to the Congo region.
In 1978/9, write Foley et al. in 2000, among the preserved serum samples of four gay men in San Francisco, there was only a 2.8% difference in nucleotides within the env gene;
while env variants collected in the United States throughout the 1980s and 1990s showed, on average, increasing genetic diversity and divergence from the subtype B consensus sequence. These results indicate that the majority of HIV-1 currently circulating in the United States may be descended from an initial introduction and rapid spread during the mid- to late 1970s of subtype B viruses with limited variability (i.e., a founder effect)3
This is to say that while the B strain of HIV-1M spread predominantly at first in the US, others later crossed over; the Congo region in other words remained a reservoir of other, pre-existing HIV-1M strains that were able to eventually take root elsewhere. As already mentioned, the earliest Congo samples were from the A and D subgroups.
Thus to the question of whether this Congo region HIV-1M reservoir attained its genetic diversity in humans or represents multiple 1M introductions from central-African-chimpanzees. The answer derives, to a great extent, from the fact that no stable HIV-1M virus exists in chimpanzees.
It has in fact been proposed by turn-of-the-millennium polio-HIV theorist Edward Hooper that the answer to phylogenetic estimates which rule out zoonotic cross-over in the 50s polio vaccine era is that HIV-1M attained its diversity in chimpanzees. This claim is answered, whether satisfactorily or not, by Sharp et al. in 2001.4
What is perhaps more conclusive than their argument, which I will not quote, is that HIV-1M is distinguished from other human HIVs by the previously mentioned adaptation of the Vpu gene to human tetherin. It is not plausible that multiple strains of a monkey virus 1M reservoir have all independently surpassed this barrier to human tropism, when none of the non-1M HIVs have managed the same. Therefore, 1M crossed into humans, adapted to tetherin, and then diversified into various subgroups — in humans.
But then why just Africa?
Still, all of this diversification of HIV-1M at least appears to have taken place and have been limited to the Congo region which is nearby the host of the ancestor to HIV-1M, i.e. central-African-chimpanzees.
If we consider that the sexual revolution and needle-based drug use were novelties of the 20th Century, nothing is really so remarkable about this apparent limitation of HIV-1M to the same geographical region of its emergence from chimpanzees. What I propose — what is my current tentative stance on the mystery — is that HIV was before modern times simply a “primate-adjacency problem” that most human cultures were removed from, simply by not being in the Congo region. The advent of gay meccas and of needle-based drug use expanded the footprint of the problem.
Nothing, further, is particularly remarkable about the lack of awareness of the problem in classical medical literature in Africa before modern times, given that the region was and still remains replete with far more obvious diseases than AIDS. How, in fact, could AIDS ever be noticed in a landscape which routinely kills even the healthy via yellow fever, etc.? It is only when HIV-1M spread (due to homosexual promiscuity and to drug use) to regions not surrounded by such lethal diseases, but rather doused in hygiene and antibiotics, that it could be noticed (due to enabling more bizarre illnesses).
Still, is human HIV truly ancient?
This leads to what is at present an ambivalent position on my part. Are Worobey, et al., correct in 2008 that HIV-1M must have emerged in humans in the early 20th Century, or not?
The problem in answering this question is that even though the pre-existing diversity of HIV-1M is undeniable, the spread of this diversity beyond the Congo region remains obviously “novel.” It would seem that a truly ancient human-adapted retrovirus should have been provably present everywhere, and particularly in regions that could have noticed the disease conditions that require hygiene to perceive — what could stop it from being so?
Thus, I grant that their dating of HIV-1M’s most recent common ancestor to the early 20th Century is prima facie consistent with the apparent limitation of the virus to the Congo region before the 1980s.
That said, major caveats ought to be mentioned.
To begin with, phylogenetic estimates of most recent common ancestors of viruses are only the floor, not the ceiling, of the possible introduction of the same virus into the host niche (even though they are treated, explicitly, as ceilings). That molecular clock algorithms think HIV-1M subgroups diverged around the turn of the 20th Century simply means that any HIV-1M in humans today derived from a version in humans around that time; it does not mean that HIV-1M sprang into existence around that date. Currently, all circulating versions of SARS-CoV-2 derive from Omicron; but Omicron derives from a virus that was circulating over 20 months before it was detected. Therefore, the last common ancestor of the Omicron variants is not by any means the marker of when SARS-CoV-2 crossed into human circulation. This is to say, even if phylogenetic and molecular clock algorithms were instruments of divine precision, they would still not measure and report the genetic and temporal footprint of host viruses that have been lost to time. They cannot see what is not sequenced.
In this sense, all that Worobey, et al.’s flagstone for HIV-1M’s crossover represents is a lowest “without going over” bid for the virus’s emergence into humans. Nothing that distinguishes this bid from older crossovers is available to our present base of evidence; but that may simply reflect that older and more divergent strains of HIV-1M, perhaps as old as humanity itself, have failed to stay alive. This wouldn’t be alarming in any respect.
So, all we know is that the subgroups of HIV-1M “in play” after the 1950s were probably the progeny of a single genome floating around about a half-century before.
A further problem with the idea that HIV-1M spilled into humans recently is that, well, humans have rendered the ancestral host of HIV — central-African chimpanzees — endangered. Again, I will seek an ambivalent position on this point. Yes, the late 20th Century saw an expansion in the use of primates for medical research, but it also saw near-elimination of many primate species. Thus it is said of the hosts of the closest relative of HIV,
Of the many primate lentiviruses that have been identified, SIVcpz has been of particular interest because of its close genetic relationship to HIV-1 (Fig. 2). However, studies of this virus have proven to be challenging because of the endangered status of chimpanzees.
The first isolates of SIVcpz were all derived from animals housed in primate centers or sanctuaries, although infection was rare in these populations. Collective analyses of nearly 2,000 wild-caught or captive-born apes identified fewer than a dozen SIVcpz positive individuals5
The “default narrative” of the HIV pandemic is that humans caught the virus from centrail-African chimpanzees in the same era as the same animal was being reduced to endangered status by human development, rather than that the virus had transferred long ago and was already adapted to humans living in Africa. As frequently occurs, this is founded upon a nebulous concept of information-age tropical humans now admixing with animals in some way that did not occur before modern times. In other words, it is held that tropical humans used to live next to animals all the time, but never thought of interacting with them until the vague influence of combustible engines and TVs. Why? Who knows.
Finally, it must be noted that the number of subgroups of HIV-1M is very similar to that of endemic primate SIVs — and, has not increased much in humans since being first elucidated. How could the global “pandemic” fail so notably to produce new subgroups, if the virus were truly novel?
Instead, what has been observed is that the post-80s global proliferation of HIV infections has produced a subsequent proliferation of recombinations of these same subgroups. This, I would argue, is evidence that before the 1980s, HIV-1M evolution was largely free of the influence of super-African sexual and drug-related transmission, so that subgroups rarely encountered each other within the same infected cell — and therefore that any estimates of last common ancestry based on observed mutation rates is wildly off-base, due to inferring the past from an unnatural status quo.
But overall, again, my stance at present is ambivalent. Maybe HIV-1M solved the evolutionary barrier of suppressing human tetherin around the turn of the 20th Century, maybe it did so a million years ago. If in fact it was predominately latent in Congo region cities at the beginning of the cell culture and gene sequencing era, it still remains an interesting question whether the Hepatitis B vaccine had a substantial role in causing the recognized AIDS pandemic.
While Pt. 2 will answer this question, I have written a follow-up “Pt. 1.5” which largely repeats and expands the summary arguments offered here.

{kind=link}
If you derived value from this post, please drop a few coins in your fact-barista’s tip jar.


Of course, one might propose that the Hebrew-Christian myth of Sodom and Gomorrah may have as origin ancient HIV outbreaks — however, there is not in fact any basis in either the Old or New Testament for inferring that homosexuality was going on in these places, and presumably it would have been mentioned outright if so. The association of homosexuality with these myths is merely an apocryphal understanding of the text according to Anglosphere sexual mores.
Worobey, M. et al. “Direct evidence of extensive diversity of HIV-1 in Kinshasa by 1960.” Nature. 2008 Oct 2;455(7213):661-4.
Foley, B. Pan, H. Buchbinder, S. Delwart, EL. “Apparent founder effect during the early years of the San Francisco HIV type 1 epidemic (1978-1979)” AIDS Res Hum Retroviruses. 2000 Oct 10;16(15):1463-9.
Sharp, PM. et al. “The origins of acquired immune deficiency syndrome viruses: where and when?” Philos Trans R Soc Lond B Biol Sci. 2001 Jun 29; 356(1410): 867–876.
Sharp, PM. Hahn, BH. “Origins of HIV and the AIDS Pandemic.” Cold Spring Harb Perspect Med. 2011 Sep; 1(1): a006841.
All of the evidence which I’ve examined and considered argues strongly that the virus referred to as HIV is a rather widespread and benign retrovirus which is utterly incapable of creating a life-threatening immune system depression in anyone whose immune system is healthy, properly nutrified, and functioning competently.
The AIDS scare principally involved three groups: 1) highly promiscuous gay males, 2) intravenous drug abusers, and 3) hemophiliacs. These seemingly disparate groups had one very important feature in common - the chronic and excessive use of chemicals which were toxic to the progenitor cells needed to maintain healthy levels of immune cell populations. Many of these mother cells are located in the marrow of the long bones because they're extremely sensitive to background radiation. So, nature, in her wisdom, saw fit to bury them within a densely calcified matrix. However, they're equally sensitive to chemical toxins, as seen with cancer chemotherapy patients who won't be treated when their T and B cell counts are still too low. I’ll repeat that with emphasis, STILL TOO LOW. That’s clinic-speak for “that last burst of poisonous chemicals which we pumped into you wreaked havoc on your sensitive stem cells and they’re struggling to make a comeback”.
Now, the "fast-track" gay males were as promiscuous in their use of recreational drugs as they were in their pursuit of virtually anonymous sexual expression. Poppers, for example, are extremely toxic to stem cells of all types. Chronic use is a virtual prescription for a dangerously under-performing immune system.
The intravenous drug abusers were not only using copious amounts of highly concentrated and immune-compromising recreational drugs but they were also sharing needles. Needless to say, the sharing of needles is an excellent method for the direct transmission of not only potential pathogens but also relatively harmless retroviruses such as HIV.
Lastly, the treatments for hemophiliacs involved taking clotting factors which had to be pooled from large numbers of blood donations. Consequently, it’s not at all surprising that virtually all hemophiliacs were subsequently found to have antibodies against the HIV virus.... but absolutely NO circulating titers of the virus itself.
In addition, a majority of the “AIDS" victims probably had chronic nutritional deficiencies and most engaged in practices which are known to spread parasites of every description. Consequently, most could be expected to be antibody positive for a virus which was already a ubiquitous, albeit yet to be discovered, viral parasite of humans. Unfortunately, improved techniques for the detection of unknown viruses in humans and other animals came along roughly at the same time that virologists were desperately searching to find a microbial explanation for this horrific immune deficiency syndrome, later referred to as "AIDS".
When a high percentage of people amongst all three of these at-risk groups were found to be antibody-positive for HIV, this hapless "new” virus was given a reputation which was totally undeserved. It was chronic chemical intoxication and corruption of blood that doomed those poor souls, not HIV. Any bloom of pathogenic microbes in mammals is a symptom of an underlying immune system dysfunction, PERIOD. The immune compromise occurs BEFORE the bloom of viruses, or other opportunistic infections, or exotic cancers such as Kaposi’s sarcoma, or any other benign or malignant cellular dysplasias which occur in humans at absurdly high rates.
What’s more, the “HIV causes AIDS” theory could not explain the large number of people who exhibited all of the symptoms of this new plague but who were found to have no circulating antibodies against the HIV virus. Instead of trying to explain this troubling phenomenon, the medical industrial complex chose to ignore it for political and monetary interests. Money subsequently poured into BigPharma to synthesize even more toxic drugs to treat the harmless HIV virus instead of educational programs which would instruct all citizens on how to keep their immune systems vigorously healthy.
It appears from all of this that AIDS was misnamed. It was not ACQUIRED as a severe infective agent which then spread through intimate human contact. If anything was acquired, it was the shared drugs and risky lifestyles amongst friends and lovers that were so dangerously unhealthy with regard to vigorous immune surveillance and functioning.
BTW, Dr. Peter Duesberg's book "Inventing the AIDS Virus" is essential reading for anyone who needs a far more thorough explanation for this heretical "theory" regarding what I'll always refer to as IDS, not AIDS...
> The association of homosexuality with these myths is merely an apocryphal understanding of the text according to Anglosphere sexual mores.
The Koran explicitly refers to homosexuality in its recounting of the story of Sodom and Gomorrah, and it was written ~1300 years ago, outside of the Anglosphere.
“Do you approach males among the worlds, and leave those whom your Lord has created for you as your wives? Nay, you are a people of transgressing!” - 26:165-166.